На эту статью можно ссылаться, ее адрес в интернете: www.biophys.ru/archive/congress2006/pro-p121.htm
Тиолдисульфидная система и биоритмы
В.В.Соколовский
Санкт-Петербург, Институт аналитического приборостроения РАН
Механизмы регуляции биоритмов представляют собой одну из интригующих проблем хронобиологии. В рамках этой проблемы большое значение имеет вопрос о роли внешних «задатчиков» ритмов, об их природе и механизме действия на живые организмы [Ф.И.Комаров, 1989; А.М.Алпатов, 2000]. Информация на эту тему, почерпнутая из литературы разных лет и направлений, и результаты наших многолетних исследований [В.В.Соколовский и соав., 1976, 1979, 1982а, 1982б, 1984, 1986, 1987, 1988, 1992, 1993, 1996, 2000, 2003, 2004а, 2004б] заставляют обратить внимание на некоторые существенные факты, обсуждение которых явилось целью этой статьи.
1. Тиолдисульфидная система в механизмах биорегуляции.
Среди биологически активных веществ, специфичных для химического состава живых организмов, пристальное внимание привлекают тиоловые соединения, многостороннее участие которых в механизмах жизнедеятельности становится все более очевидным.
На протяжении многих лет в литературе высказывалось мнение о «фундаментальном», «ключевом» значении тиолов в биологических явлениях и процессах [E.Barron, 1951; P.Yocelin, 1972; M.Friedman, 1973; L.Packer, 1995]
Общей особенностью тиолсодержащих веществ (биотиолов) является присутствие в их молекулах сульфгидрильных SH-групп, источником которых в большинстве случаев служит аминокислота цистеин. Размеры и масса молекул биотиолов варьируют в очень широких пределах, а их виды, физико-химические свойства и биологическая активность отличаются поразительным разнообразием. К числу биотиолов относится большая группа веществ с относительно небольшими размерами молекул, таких как цистеин и гомоцистеин, эрготионеин, липоевая кислота, кофермент А., трипептид глютатион, однако многостороннее участие тиоловых соединений в явлениях жизнедеятельности связано, главным образом, с белками. Тиоловые белки принимают участие в регуляции ферментативной активности, проницаемости клеточных мембран, клеточного деления и роста, мышечного сокращения, генерации и проведения нервного импульса, действия гормонов, транспорта биологически активных веществ, функционирования системы антиоксидантной защиты, в механизмах сигнальной передачи и других процессов.
Важным химическим свойством биотиолов, с которым непосредственно связана их специфическая биологическая активность, является высокая реакционная способность сульфгидрильных групп, обусловленная уникальными особенностями атома серы: легкой поляризуемостью электронов и подвижностью незанятых d-орбиталей, допускающей d-орбитальное замещение. [M.Friedman, 1973]. Вследствие этого биотиолы в мягких физиологических условиях легко вступают в разнообразные химические реакции (окисления, алкилирования, меркаптидообразования). Исключительное значение в биологическом плане имеют окислительно-восстановительные превращения тиолов.
Поскольку атом серы в тиолах имеет наименьшую степень окисления (-2), они проявляют сильные восстановительные свойства [В.И.Слесарев, 2000].
 Окисление тиолов слабыми окислителями
в мягких физиологических условиях приводит к образованию дисульфидов (R-S-S-R), при этом степень окисления серы
повышается до -1, а реакция является обратимой. При действии же более сильных
окислителей тиолы превращаются в производные серы (сульфокислоты и др.) со
степенью окисления серы +4 и +6, причем такое окисление практически необратимо.
Окисление тиолов слабыми окислителями
в мягких физиологических условиях приводит к образованию дисульфидов (R-S-S-R), при этом степень окисления серы
повышается до -1, а реакция является обратимой. При действии же более сильных
окислителей тиолы превращаются в производные серы (сульфокислоты и др.) со
степенью окисления серы +4 и +6, причем такое окисление практически необратимо.
Необходимо подчеркнуть, что реакционная способность SH-групп вообще и их склонность к окислению в частности зависят от особенностей микроокружения в молекуле: так, например, знак и величина электрического заряда соседних радикалов и функциональных групп увеличивают или уменьшают реактивность SH-групп, а гидрофильный или гидрофобный характер окружения облегчает или затрудняет доступность тиоловой группы для реагента. Различная реакционная способность неодинаковых SH-групп дает возможность специфически воздействовать на различные клеточные функции [А.Сент-Дьердьи, 1971].
На процесс образования дисульфидной связи в мягких окислительных условиях существенное влияние оказывает также пространственная близость или разобщенность SH-групп [E.Barron, 1951; M.Friedman, 1973]. Это обстоятельство играет особую роль в механизме формирования внутримолекулярных и межмолекулярных дисульфидных связей в белках с их подвижной пространственной структурой (конформацией). На реакционной способности тиоловых групп белка сказывается любое воздействие, способное вызвать конформационные изменения. Модификация укладки полипептидной цепи в трехмерном пространстве сопровождается изменениями состава функциональных групп аминокислотных остатков, расположенных возле SH-групп, и их влияния на ионизацию и реактивность тиоловых центров. В свою очередь, химическая модификация SH-групп белка приводит к его конформационным изменениям.
Феномен образования дисульфидной связи в результате мягкого окисления двух тиоловых групп, и последующей регенерации этих SH-групп в результате восстановительного расщепления дисульфидной связи следует рассматривать, как свидетельство обратимости редокс превращений тиолов и дисульфидов, т.е. существования единой окислительно-восстановительной тиолдисульфидной системы (ТДС), функционирование которой должно сопровождаться колебаниями тиолдисульфидного равновесия (рис. 1).
К настоящему времени в представлениях о механизмах биологической регуляции сформировалась концепция, позволяющая объединить в единое целое множество известных ранее и описанных в последние годы фактов, рассматривая их под одним общим углом зрения. Речь идет о существовании прямой зависимости биохимических реакций и связанных с ними физиологических процессов от окислительно-восстановительного состояния («redox status», ROS) биологической системы и окружающей ее среды, причем в рамках этой концепции особое внимание уделяется редокс состоянию тиолдисульфидной системы [Y.Suzuki, L.Packer, 1995; C.Sen, 1998; L.Moran, 2001; R.Daynes et al., 2003; H.Nakamura, 2004]. Накоплено множество данных о роли колебаний окислительно-восстановительного состояния ТДС клеток и тканей живых организмов в механизмах регуляции их функциональной активности [В.А.Труфанов и соав., 1990; L.Packer, 1995; C.Sen, 1998; L.Moran et al., 2001]. Вариации редокс состояния тиолдисульфидного звена нативного белка приводят к его конформационным изменениям, следствием которых является модуляция специфической биологической активности белка [H.Gilbert, 1995; L.Weiner, 1995]. К числу редоксзависимых тиоловых белков относятся многие ферменты (оксидоредуктазы, трансферазы, гидролазы и др.), белки клеточных мембран, сократительные белки мышечной ткани, белки клеточного деления и генной экспрессии, рецепторные и другие сигнальные белки, а также многие белки с иными специфическими функциями (альбумины, глобулины, фибриноген, гемоглобин и др.) [C.Padgett, A.Whorton, 1995; Y.Suzuki, L.Packer, 1995; J.Navarro et al., 1999; K.Webster et al., 2001; Y.Liu, D.Gutterman, 2002; R.Daynes et al., 2003].
Все эти данные подтверждают ранее высказанное утверждение [В.В.Соколовский, 1996, 2004а], что многочисленные и разнообразные по своей природе химические, физические и биологические агенты, способные модифицировать тиолдисульфидную систему, оказывают, тем самым, прямое влияние на биохимические и физиологические процессы, зависящие от ее редокс состояния.
Таким образом, основу тиолдисульфидного механизма биорегуляции составляет последовательность взаимодействий:
|
Внешний агент |
|
ТД редокс система |
|
конформация |
|
белка |
|
специфическая активность белка |
||
Прямым следствием обратимости реакции окисления SH-групп биотиолов при многократном повторении цикла тиолдисульфидных превращений является возможность функционирования механизма регуляции ритмов колебаний конформационных параметров и биологической активности белков, ритмов их чувствительности к внешним воздействиям.
2. Тиолдисульфидная система в биохимическом механизме действия космофизических факторов.
Еще в середине минувшего века Х.С.Коштоянц [1951] отмечал способность ряда белков отвечать на воздействие различных химических и физических факторов окружающей среды структурными изменениями молекул, подчеркивая при этом обратимый характер таких изменений. Последнее, по его мнению, «означает, что белок, претерпев структурные изменения, возвращается в первоначальное структурное состояние, что позволяет ему вновь реагировать на повторное воздействие». При этом автор связывал упомянутые свойства белка с уникальными физико-химическими свойствами SH-групп, обладающих наиболее высокой реакционной способностью и легко вступающих в нормальных физиологических условиях в обратимую окислительно-восстановительную реакцию.
Четверть века спустя те же мысли высказывает Б.А.Ройтруб [1975], а В.Я.Александров [1975] предлагает гипотезу, согласно которой «в клетках имеется какой-то регуляторный механизм или механизмы, которые в зависимости от обстоятельств могут менять уровень конформационной гибкости клеточных белков в обе стороны, повышать или снижать его», что является важной стороной процесса адаптации.
Последние годы внесли более уверенные интонации и конкретное содержание в соответствующие формулировки. Так, J.Hancock et al. [2004] считают, что окисление или восстановление цистеиновых остатков в молекулах белков ведет к нарушению конформации белков и, соответственно, к модуляции их функций. Теперь – отмечают Y.Rahman et al.[2005] – стало известно, что многие белки содержат высоко реакционноспособные цистеиновые фрагменты, локализованные в активных регуляторных центрах, которые являются первичными мишенями окислительных модификаций и таким образом важными компонентами механизма редокс сигнализации. Однако механизм, посредством которого оксиданты в действительности участвуют в редокс сигнализации, по мнению авторов, еще нуждается в разъяснении. Все вышесказанное означает, что зависимость конформации и функциональной активности белка от его редокс состояния признается в настоящее время несомненным фактом, а обратимость редокс превращений тиолдисульфидной системы воспринимается как аксиома уже давно. Все это в целом подтверждает сформулированный выше вывод о существовании окислительно-восстановительного механизма регуляции биоритмов, в котором ведущее значение имеет тиолдисульфидное звено.
Некоторые виды таких ритмов уже описаны в литературе. Так, ритмические изменения концентрации SH и –S-S- групп в клетках живых организмов были обнаружены при изучении процессов клеточного деления [И.А.Алов, 1972; S.Pandeya, 1979] и функционирования контрактильных белков [H.Sakai, 1965]. Были выявлены колебания величины тиолдисульфидного отношения (ТДО) в белках печени, происходящие в течение суток, установлено сходство ритмов этих колебаний во фракциях клеточных ядер, микросом и цитозоля, и их корреляция с суточными вариациями активности тиолтрансферазы, катализирующей образование смешанного дисульфида «глютатион-белок» [Y.Jsaaks, F.Binkley, 1977]. В нашей лаборатории обнаружены значительные различия величин ТДО при их измерении в разные периоды суток в белковой и не содержащей белков фракциях крови белых крыс [Г.А.Баскович и соав., 1986].
Наблюдение «спонтанных» колебаний концентрации SH-групп и одновременных колебаний активности тиолзависимого фермента аденозинтрифосфатазы в растворах мышечных белков актина, миозина и актомиозина (in vitro) [С.Э.Шноль и соав., 1964, 1967, 1979], установление сильной зависимости амплитуды конформационных колебаний белков актомиозинового комплекса от концентрации в инкубационной среде окислителей и восстановителей привели С.Э.Шноля к важному выводу о том, что вероятной причиной колебаний концентрации SH-групп являются реакции «обратимого взаимопревращения сульфгидрильных и дисульфидных групп» [С.Э.Шноль, 1967]. В последующее время выявление сходства флуктуаций биохимических тестов и других тестовых показателей (измерения радиоактивности, электрофоретической подвижности клеток и др.) послужило основанием для предположения, что макрофлуктуации скоростей различных биохимических и химических реакций (окисление аскорбиновой кислоты дихлорфенолиндофенолом), по-видимому, обусловлены космофизическими причинами [Н.В.Удальцова, В.А.Коломбет, С.Э.Шноль, 1987]. Результаты наших многолетних исследований, имевших целью изучение биохимического механизма реакции живых организмов на действие космофизических факторов (КФФ), приводимые ниже, оказались в своей основе созвучны этим соображениям.
В свое время вопрос о том, не является ли тиолдисульфидная система «мишенью» диоксида азота в механизме токсического действия последнего поставил нас перед необходимостью выбора модели тиола для проведения экспериментов in vitro. С этой целью был использован «унитиол» (димеркаптопропансульфонат натрия), в молекуле которого присутствуют две SH-группы, расположенные в максимальной близости одна от другой, что в сильной степени облегчает образование внутримолекулярной дисульфидной связи при их окислении в мягких условиях [Б.И.Гольдштейн, 1951]. Таким образом, модель позволяет имитировать функциональный тиоловый центр многих белков, ответственный за их редокс превращения и необходимый для регуляции их специфической биологической активности.
|
унитиол: |
|
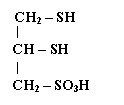
В серии опытов измеряли скорость реакции окисления унитиола нитритным ионом [В.В.Соколовский, Р.Н.Павлова, 1976; В.В.Соколовский, 1982]. Смесь водных растворов унитиола и нитрита натрия (фосфатный буфер, pH 7,0) инкубировали в ультратермостате при температуре 39,00С в течение 2 часов, определяя каждые 30 мин. остаточную концентрацию SH-групп (амперометрическим титрованием нитратом серебра). Графическим путем определяли время, необходимое для окисления 50% SH-групп реакционной смеси, т.е. «время полуокисления» [E.Barron, 1951] унитиола (ВПОУ). Как уже говорилось, на скорость взаимодействия SH-групп с окислителем оказывают влияние многие факторы, такие как химическая природа тиола и окисляющего вещества, их стехиометрические соотношения, температура и pH среды, ионная сила раствора. При постоянстве этих условий, казалось бы, скорость окисления тиола в повторяющихся наблюдениях должна оставаться одинаковой. Между тем, оказалось, что величина ВПОУ, повторно измеряемая в однозначных условиях в течение продолжительного времени, претерпевала разнонаправленные изменения. В отдельные дни одного или даже нескольких месяцев она оставалась сравнительно стабильной или варьировала в узком интервале времени, тогда как в другие периоды ее колебания достигали значительного размаха. Убедившись, что такие эпизоды не связаны с методическими погрешностями, мы допустили возможность влияния некоего внешнего фактора на кинетику реакции и сделали попытку сопоставить полученные данные с уровнем солнечной активности (СА). Удовлетворительную корреляцию ВПОУ с числами Вольфа, характеризующими относительное количество пятен на видимом солнечном диске, обнаружить не удалось. Вместе с тем, сведения, публикуемые Пулковской обсерваторией в бюллетене «Солнечные данные», свидетельствовали, что 20 и 22.01.1971, в дни, когда мы наблюдали значительное уменьшение ВПОУ (т.е. увеличение скорости реакции) происходило многократное увеличение (против обычного уровня) потоков радиоизлучения Солнца в диапазонах 204 и 234 МГЦ (20.01.1971), наблюдались и другие необычные явления: хромосферные вспышки с несколькими максимумами яркости (22 и 23.01), появление очень протяженной активной области (21 и 23.01). В другом случае резкого уменьшения ВПОУ (22.04.1972) значительное усиление потока радиоизлучений в диапазоне 100 МГц было отмечено 21 и 22.04., причем 21.04. в метровом диапазоне радиоволн наблюдался необычный всплеск продолжительностью 210 минут.
Интересные факты были обнаружены в
нашей лаборатории В.М.Тимофеевой в 1970-
|
|
Рис.2. ВПО унитиола адренохромом в декабре |
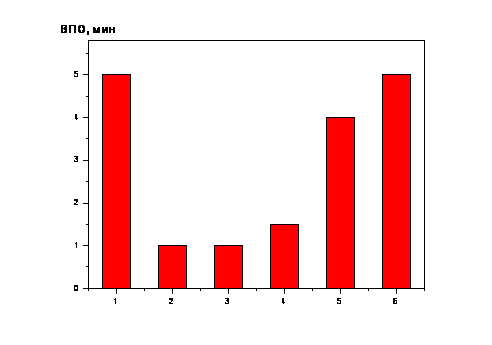
Еще один случай ускорения окислительной модификации унитиола адренохромом, связанный с кратковременным возмущением солнечной активности, был отмечен 20.04.1971г., когда ВПОУ составило всего 15 сек. при обычном («фоновом») уровне порядка 5 минут. Своеобразие этого эпизода состояло в быстрой нормализации ВПО, величина которого уже на следующие сутки равнялась 6,5 мин. При этом вспышечные индексы 20-го и 21.04. резко различались между собой: 277 и 14 соответственно. Другой особенностью эпизода явилось проявление 21.04. геомагнитной возмущенности, индекс которой (Ар) возрос с 4 (20.04.) до 22 (21.04.) [Solar Geofis. Date, 1971; Проблемы космической биологии, 1982]. Этот факт позволяет видеть, что именно всплеск солнечной активности, а не геомагнитное возмущение, послужил толчком к ускорению окислительной реакции.
С целью дальнейшего изучения феномена существования связи между скоростью реакции окисления тиоловых соединений в растворах (in vitro) и состоянием солнечной активности проводили ежедневные (за исключением выходных дней) измерения ВПОУ нитритным ионом в течение 34 месяцев (период с сентября 1974г. по июнь 1977г.). При этом неизменно соблюдали стандартные условия опытов и время их проведения (10-12 часов). Для характеристики уровня солнечной активности (СА) использовали следующие показатели: числа Вольфа (W); величину интенсивности потоков радиоизлучений (R) с частотой 200 МГц; суммарное число хромосферных вспышек (N) за сутки. Кривые, иллюстрирующие динамику вариаций среднемесячных величин ВПОУ и индексов СА за период наблюдений, приведены на рис.3. Для устранения небольших колебаний предпринято сглаживание рядов по трем точкам по формуле невзвешенной скользящей средней [А.Л.Чижевский, 1976]. Для количественной оценки тесноты связи между отдельными показателями использовали метод вычисления коэффициента линейной корреляции Пирсона.
|
|
Рис.3. Динамика вариаций среднемесячных величин ВПОУ и индексов СА за период наблюдений с сентября 1974г. по июнь 1977г. |
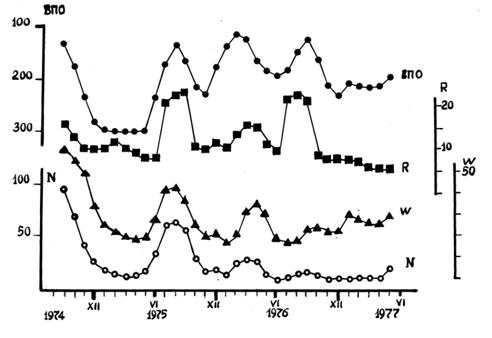
Кривая ВПОУ на рис.3 позволяет
видеть, что средняя скорость реакции в различные месяцы то уменьшалась, то
увеличивалась, причем за весь период наблюдений сильное уменьшение ВПОУ (т.е. ускорение
процесса окисления) происходило 4 раза. Уровень солнечной активности тоже
изменялся и имел 4 синхронных максимума. Это обстоятельство в свою очередь
указывает на существование связи механизма регуляции скорости окислительной
реакции с физическими явлениями на Солнце – связи, проявляющейся и в достаточно
продолжительных наблюдениях. Обращает на себя внимание участок кривой ВПОУ,
соответствующий 1974-
Общее сходство конфигурации
кривых, представленных на рис.3, наиболее отчетливо проявлялось в период,
соответствующий 21 месяцу наблюдений с момента их начала. Этот факт заставляет
думать о происходивших в течение 34 месяцев вариациях тесноты связи между
величинами ВПОУ и индексов, использованных для количественной оценки СА. О том
же свидетельствуют и результаты анализа коэффициентов корреляции. Обратная связь
ВПОУ с числом вспышек и интенсивностью потока радиоизлучений Солнца оказалась в
1974-
Вместе с тем, реакцию окисления унитиола нитритом следует рассматривать как отдельный пример медленных окислительных реакций, на скорость которых оказывает влияние солнечная активность. Выше уже говорилось о реакции окисления унитиола адренохромом. Связь с уровнем СА проявилась также при измерениях скорости окисления красителя «основной синий К» гидропероксидом [В.А.Кулеба, В.А.Бандюкова, 1979] и окисления унитиола нингидрином [Р.Н.Павлова, В.С.Сорокина, 1986]. К числу таких реакций надо отнести и реакцию полимеризации акрилонитрила, высоко чувствительную к действию свободных радикалов и составляющую основу одного из химических тестов, предложенных Дж.Пиккарди для изучения эффектов СА [Дж.Пиккарди, 1967; Р.Н.Павлова, Н.К.Зайцева, 1984].
Как уже говорилось, обратимое окисление сульфгидрильных групп делает возможным функционирование тиолдисульфидного редокс механизма биологической регуляции. Зависимость скорости окисления тиолов от уровня СА, обнаруженная в опытах с водными растворами (in vitro), наводит на мысль о существовании зависимости редокс состояния живых организмов от космофизических воздействий и о возможности космофизической регуляции процессов жизнедеятельности путем модификации ТДС. Уверенность, что воздействие Космоса на окружающую среду и на живой организм осуществляется одновременно, что внешний и внутренний мир реагируют совместно на основные явления, происходящие в Космосе, высказывал Дж.Пиккарди [1967]. В связи с этими соображениями нами были предприняты поиски свидетельств участия тиоловых соединений в механизмах реакции живых организмов на действие космофизических факторов (in vivo). Увеличение скорости окисления тиоловых соединений при усилении вспышечной деятельности Солнца (или связанное с иными космофизическими явлениями) может иметь прямое отношение к молекулярным механизмам влияния этих факторов на биосферу Земли. В этом плане заслуживает внимания возможность изменения активности ферментов, каталитические свойства которых обусловлены присутствием тиоловых групп в молекуле фермента. Эти функциональные группы либо входят в состав активного центра фермента, либо ответственны за поддержание каталитически активной конформации белковой молекулы.
Хорошо известно, что типичным представителем тиоловых ферментов являются аденозинтрифосфатазы (АТФазы), которые легко и обратимо ингибируются при действии слабых окислителей (и тяжелых металлов) и реактивируются в присутствии тиоловых соединений. Эти ферменты катализируют гидролитическое расщепление аденозинтрифосфата (АТФ) и играют универсальную роль в процессах жизнедеятельности (энергетический обмен, трансмембранный перенос катионов, мышечное сокращение, генерация и проведение нервного импульса и др.).
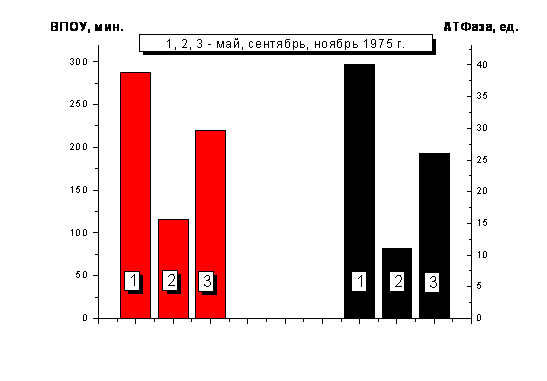
В связи с этими соображениями
определяли общую активность АТФаз головного мозга белых крыс в разные периоды
времени, для которых была характерна либо низкая, либо высокая скорость
окисления унитиола (т.е. соответственно, большая или малая величина ВПОУ).
Опыты были проведены в мае, сентябре и ноябре 1975г. Активность АТФаз оценивали
по скорости расщепления АТФ, выражая ее в мкг неорганического фосфора (Рн)
на 1 мг белка в час. Результаты оказались следующими. Если средняя скорость
окисления унитиола в дни наблюдений в мае (12 набл.) была низкой (ВПОУ=290![]() 10
мин.), то в сентябре (16 набл.) она значительно повысилась (ВПОУ=118
10
мин.), то в сентябре (16 набл.) она значительно повысилась (ВПОУ=118![]() 26
мин.), а в ноябре (8 набл.) вновь уменьшилась (ВПОУ=218
26
мин.), а в ноябре (8 набл.) вновь уменьшилась (ВПОУ=218![]() 33
мин.). При этом АТФазная активность нервной ткани, высокая в мае (39,5
33
мин.). При этом АТФазная активность нервной ткани, высокая в мае (39,5![]() 4
ед.), резко упала в сентябре (10,7
4
ед.), резко упала в сентябре (10,7![]() 0,8 ед.), а в ноябре
увеличилась до 26,7
0,8 ед.), а в ноябре
увеличилась до 26,7![]() 13,9 ед. (рис.4). Та же закономерность
проявилась в опытах с определением АТФазной активности мембран эритроцитов
барана в июне и сентябре 1975г., когда коэффициент корреляции между активностью
фермента в клетках крови и величиной ВПОУ равнялся 0,87 (при р=0,05). Таким
образом, обнаруживается существование определенной связи активности АТФазы
нервной ткани со скоростью окисления унитиола in vitro, что можно рассматривать как
признак участия одного из жизненно важных тиоловых белков в реакции живого
организма на внешнее воздействие, имеющее, по всей вероятности, космофизическое
происхождение.
13,9 ед. (рис.4). Та же закономерность
проявилась в опытах с определением АТФазной активности мембран эритроцитов
барана в июне и сентябре 1975г., когда коэффициент корреляции между активностью
фермента в клетках крови и величиной ВПОУ равнялся 0,87 (при р=0,05). Таким
образом, обнаруживается существование определенной связи активности АТФазы
нервной ткани со скоростью окисления унитиола in vitro, что можно рассматривать как
признак участия одного из жизненно важных тиоловых белков в реакции живого
организма на внешнее воздействие, имеющее, по всей вероятности, космофизическое
происхождение.
|
|
Рис.4. АТФазная активность ткани головного мозга белых крыс и
ВПОУ в |
В качестве наиболее вероятного
источника такого воздействия в приведенных выше случаях мы рассматривали активное
Солнце. Однако в 1982-
Многократные ежедневные измерения ВПОУ проводились в период глубокого минимума СА с 15.07.1996г. по 01.06.1997г. в экологически чистых условиях Антарктики, на полярной станции «Мирный» [Э.С.Горшков, С.Н.Шаповалов]. Анализ полученных в этих наблюдениях данных позволил обнаружить связь общей динамики изменений скорости тестовой реакции (окисления унитиола нитритом) с вариациями гравитационного поля, обусловленными поступательным и вращательным движением Земли, особенностями взаимодействия масс Солнца, Земли и Луны. В спектре изменений ВПОУ отчетливо проявились периоды, близкие к периодам основных возмущений от Солнца: 31,8 сут.; 14,8 сут. [Э.С.Горшков и соав., 2000, 2004]. При последующем анализе оказалось, что значительные флуктуации ВПОУ на протяжении 21-го солнечного цикла были наиболее тесно связаны с одним из двух доминирующих факторов: в период максимальной солнечной активности эти флуктуации были обусловлены преимущественно ее проявлениями, тогда как в периоды минимальной активности они определялись главным образом гравитационными возмущениями от Солнца. Все эти данные детально рассматриваются в статье [O.A.Troshichev et al., 2004]. Здесь же уместно подчеркнуть, что их получение позволило включить гравитационное поле в число космофизических агентов, с ритмикой которых коррелируют скорость реакции окисления унитиола нитритом in vitro и ряд иных показателей.
Так, в серии исследований,
выполненных в 2001-
Были, в частности, выявлены суточные колебания активности тиолзависимого фермента моноаминоксидазы в плазме крови кроликов [Р.Н.Павлова и соав., 1984]; суточные вариации ВПОУ и тиолдисульфидного отношения крови белых крыс [Г.А.Баскович и соав., 1986]; возрастание ВПОУ при наступлении полярного дня [Э.С.Горшков и соав., 1997]; колебания скорости реакции окисления аскорбиновой кислоты дихлорфенолиндофенолом в разное время суток [Н.В.Удальцова и соав., 1987], а также в периоды новолуний, полнолуний и солнечных затмений [А.К.Кириллов, 2003].
Таким образом, итогом наших многолетних исследований стало обнаружение признаков влияния космофизических факторов на скорость окисления унитиола в водном растворе и на редокс состояние тиоловых соединений в живых организмах.
Эти данные, суммированные в настоящей работе и рассмотренные в свете представлений о сенсорнорегуляторных функциях тиолдисульфидной системы, способствуют пониманию биохимического механизма реакции живых организмов на флуктуации солнечной активности и околоземного гравитационного поля. Вызванное этими факторами ускорение или замедление окисления SH-групп сопровождается изменением концентрации восстановленных и окисленных тиолов и, соответственно, - величины тиолдисульфидного отношения. Изменение редокс состояния тиолдисульфидной системы, в свою очередь, приводит к модификации пространственной структуры белков и модуляции их специфической биологической активности.
Полученные данные позволяют утверждать, что флуктуации солнечной активности и гравитационного поля могут оказывать постоянное действие на биологические ритмы, обусловленные состоянием тиолдисульфидной системы.
Резюме.
В настоящей работе суммированы итоги многолетних исследований влияния космофизических факторов на скорость реакции окисления унитиола в водном растворе и на редокс состояние тиоловых соединений в живых организмах. Результаты работы, рассмотренные в свете представлений о сенсорно-регуляторных функциях тиолдисульфидной системы, способствуют пониманию биохимического механизма реакции живых организмов на ритмы флуктуаций солнечной активности и околоземного гравитационного поля.
Ускорение или замедление окисления SH-групп сопровождается изменением концентрации восстановленных и окисленных форм тиолов и, соответственно, - величины тиолдисульфидного отношения. Изменение редокс состояния тиолдисульфидной системы, в свою очередь, приводит к модификации пространственной структуры белков и модуляции их специфической биологической активности.
Литература:
1. Barron E. – Thiol groups of biological importance. // Adv. enzymol., 1951, v. 11, pp. 201-266.
2. Jocelyn P. – Biochemistry of the SH-group. The occurrence, chemical properties, metabolism and biological function of thiols and disulphides. – London – N. – Y; - Ac. Press, 1972. – 404 pp.
3. Friedman M. – Chemistry and biochemistry of sulfhydryl group in amino acids, peptides and proteins. – Oxford, Perg. Press, 1973. – 448 pp.
4. Packer L. (ed.) – Biothiols. Part A: Monothiols and dithiols, protein thiols and thiyl radicals. // Methods in Enzymol. – San Diego. – Ac. Press – 1995, vol. 251. – 382 pp.
5. Слесарев В.И. – Химия. Основы химии живого. СПб. – Химиздат. 2000. – 767с.
6. Сент-Дьердьи А. – Биоэлектроника. – «Мир» – М. – 1971. 79с.
7. Suzuki J., Packer L. – Redox regulation of DNA – protein interactions by biothiols. // Methods in Enzymol. – San Diego. Acad. Press. – 1995. – v. 252. – p. 175-180.
8. Sen C. – Redox signaling and the emerging therapeutic potential of thiol antioxidant. // Biochem. Pharmacol. – 1998. – v. 55, – № 11. – p. 1747-1758.
9. Moran L., Gutteridge J., Quinlan G. – Thiols in cellular redox signaling and control. // Curr. Med. Chem., 2001. – v. 8, – №7. – p. 763-772.
10. Daynes R., Enioutina E., Jones D. – Role of redox imbalance in the molecular mechanisms responsible for immunosenescence. // Antioxid and redox signal. – 2003. – v.5, – №5. – p. 537-548.
11. Nakamura H. – Thioredoxin as a key molecule in redox signaling. // Antioxid and redox signal. – 2004. – v.6, – №1. – p. 15-17.
12. Труфанов В.А., Кичатинова С.В., Шатилов В.Р., Кретович В.Л. – Ферменты тиолдисульфидного обмена белков. // Прикл. биохим. и микробиол. – 1990. – Т.26. – №1. – с.3-10.
13. Gilbert H. – Thiol / disulfide exchange eguilibria and disulfide bond stability. // Methods in Enzymol. – San Diego. – Ac. Press. 1995. – v.251. – p. 8-28.
14. Weiner L. – Quantitative determination of thiol groups in low and high molecular weight compounds by electron paramagnetic resonance. // Methods in Enzymol. – 1995. – v.251. – p. 87-106.
15. Padgett C., Whorton A. – Regulation of cellular thiol redox status by nitric oxide. // Cell. biochem. biophys. – 1995. – v.27. - №3. – p. 157-177.
16. Navarro J., Carretero J. – Changes in glutathione status and the antioxidant system in blood and in cancer cells associate with tumour growth in vivo. // Free radic. biol. Med. – 1999. –v.26. –№3-4.–p.410-418.
17. Webster K., Prentice H., Bishopric N. – Oxidation of zinc finger transcription factors: physiological conseguences. // Antioxid and redox signal. – 2001. – v.3. – №4. – p. 535-548.
18. Liu Y., Gutterman D. – Oxidative stress and potassium channel function. // Clin. exper. pharmacol. physiol., – 2002. – v.29. – p. 305-311.
19. Коштоянц Х.С. – Белковые тела. Обмен веществ и нервная регуляция.–1951.–М.–АН СССР.100с.
20. Ройструб Б.А. – Конформационные переходы в белках крови при различных функциональных состояниях нервной системы. // «Наукова думка». – Киев. – 1975. – 188с.
21. Александров В.А. – Клетки, макромолекулы и температура. «Наука». – Л. – 1975. – 328с.
22. Hancock J., Desikan R., Neill S., Cross A. – New equations for redox and nano-signal transduction. // J.Theor. Biol., – 2004. – v.226. – №1. – p. 65-68.
23. Rahman J., Biswas K., Jimenez L., Torres M., Forman H. – Glutathione, stress responces and redox signaling in lung inflammation. // Antioxid and redox signal. – 2005. – v.7. – №1-2. – p. 42-59.
24. Алов И.А. – Цитофизиология и патология митоза. // М., Медицина. – 1972. – 264с.
25. Pandeya S. – Role of sulphydryl compounds in biological systems. // J.Scient and industry. research. – 1979. – v.38. – p. 568-593.
26. Sakai H. – Studies on sulfhydryl groups during cell division of sea urchin egg. // Biochim. et Biophys. Acta. – 1965. – v.102. – №1. – p. 235-248/
27. Isaacs J., Binkley F. – Glutathione dependent control of protein disulfide – sulfhydryl content by subcellular fractions of hepatic tissue. // Biochim. et Biophys. Acta. – 1977. – v.497. – №1. – p. 192-204.
28. Баскович Г.А., Макаров В.Г., Киселева Н.Н. – Влияние суточных ритмов на состояние тиолдисульфидной системы крови и тканей животных при вибрации и введении дибазола. // Флуктуации состояния биохимических систем. – Сб. науч. тр. ЛСГМИ. – Л. – 1986. – с. 40-44.
29. Шноль С.Э., Смирнова Н.А. – Колебания концентрации SH-групп в растворах актомиозина, актина и миозина. // Биофизика. – 1964. – т.9. – вып. 4. – с. 532-534.
30. Шноль С.Э. – Конформационные колебания макромолекул. // Колебательные процессы в биологических и химических системах. – Наука. – М. – 1967. – с. 22-26.
31. Рыбина В.В., Шноль С.Э. – Синхронные конформационные колебания титра сульфгидрильных групп в растворах белков. Обратимое окисление как возможная причина этого явления. // Биофизика. – 1979. – т.24. – вып.6. – с. 970-976.
32. Удальцова Н.В., Коломбет В.А., Шноль С.Э. – Возможная космофизическая обусловленность макроскопических флуктуаций в процессах разной природы. – АН СССР. – Пущино. – 1987. – 95 с.
33. Гольдштейн Б.И. – Тиоловые группы тканевых белков, их химические свойства и биологическое значение. // Тиоловые соединения в медицине. – Госмедиздат УССР. – Киев. – 1959. – с. 49-52.
34. Соколовский В.В., Павлова Р.Н. – Окисление тиолов и солнечная активность. // Солнце, электричество, жизнь. – МОИП. – М. – 1976. – с. 9-12.
35. Соколовский В.В. – Ускорение окисления тиоловых соединений при возрастании солнечной активности. // Проблемы космич. биол., т.43: Влияние солнечной активности на биосферу. – 1982. – Наука. – М. – с. 194-197.
36. Соколовский В.В., Павлова Р.Н. – Окисление тиолов и солнечная активность. // Солнце, электричество, жизнь. МОИП. – М. – 1976. – с. 9-12.
37. Соколовский В.В. (ред.) – Тиоловые соединения в биохимических механизмах патологических процессов. // Труды ЛСГМИ, т. 125. – Л. – 1979. – 88 с.
38. Соколовский В.В. – О биохимическом механизме реакции живых организмов на изменения солнечной активности на биосферу. – «Наука». – М. – 1982а – с. 180-193.
39. Соколовский В.В. – Ускорение окисления тиоловых соединений при возрастании солнечной активности. // Проблемы космич. биол., т.43. – «Наука». – М. – 1982б. – с. 194–197.
40. Соколовский В.В. (ред.) – Антиоксиданты и адаптация. // Сб. науч. трудов. – ЛСГМИ. – Л. – 1984. – 63с.
41. Соколовский В.В. (ред.) – Флуктуации состояния биохимических систем. // Сб. науч. трудов. – ЛСГМИ. – Л. – 1986. – 75с.
42. Соколовский В.В., Гончарова Л.Л., Киселева Н.Н., Макарова И.Н., Родионова Л.П. – Антиоксидантная система организма при шумовом стрессе. // Вопросы медиц. химии. – 1987. – т.33. - №6. – с. 111–113.
43. Соколовский В.В. – Тиоловые антиоксиданты в молекулярных механизмах неспецифической реакции организма на экстремальное воздействие. // Вопросы медиц. химии. – 1988. – т. 34. - №6. – с. 2-11.
44. Соколовский В.В., Шлейкин А.Г. – Роль окислительно – восстановительных процессов в организме при действии повреждающих факторов. // Современные проблемы изучения и сохранения биосферы. – т. 2: Живые системы под внешним воздействием. – «Гидрометеоиздат». – СПб. – 1992. – с. 132-136.
45. Соколовский В.В. – Антиоксиданты в профилактике и терапии заболеваний. // Междунар. медиц. обзоры. – 1993. – т.1. - №1. – с. 11-14.
46. Соколовский В.В. – Тиолдисульфидное соотношение крови как показатель состояния неспецифической резистентности организма. // МАПО. – СПб. – 1996. – 30с.
47. Соколовский В.В., Галль Л.Н. – Оценка состояния тиолдисульфидной системы живых организмов – одно из перспективных направлений экологических исследований. // Слабые и сверхслабые поля и излучения в биологии и медицине. – 2й Междунар. Конгресс. Труды. – СПб. – 2000. – с. 21-28.
48. Соколовский В.В., Горшков Э.С., Иванов В.В., Шаповалов С.Н., Трошичев О.А. – Проявление связи вариаций состояния редокс систем в водном растворе и в организме человека с флуктуациями гравитационного поля. // Труды 3го Междунар. Конгр. «Слабые и сверхслабые поля и излучения в биологии и медицине». – Спб. – 2003. – с. 69-73.
49. Соколовский В.В. – Тиолдисульфидная система в биохимическом механизме реакции организма на экстремальное воздействие. // Вестник СПб Гос. Медиц. Академии им. И.И.Мечникова. – 2004а. - №4(5). – с. 97-100.
50. Sokolovskii V.V., Gorshkov E.S., Ivanov V.V., Shapovalov S.N., Troshichev O.A. – Relation of the regular gravitational field variations to biochemical processes observed in vitro and in vivo // Biophysics. – 2004б. – vol. 49. Suppl. 1. – s. 85 – s 91.
51. Чижевский А.Л. – Земное эхо солнечных бурь. – «Мысль». – М. – 1976. – 366с.
52. Кулеба В.А., Бандюкова В.А. – Сравнительный анализ антиоксидантной активности биофлавоноидов методом определения времени полуокисления унитиола. // Тиоловые соединения в биохимических механизмах патологических процессов. – Труды ЛСГМИ. – Л. –1979. –т.125. – с.43-46.
53. Павлова Р.Н., Сорокина В.С. – Спонтанные флуктуации окислительно – восстановительного состояния растворов. // Флуктуации состояния биохимических систем. – Сб. науч. тр. ЛСГМИ. – Л. – 1986. – с. 5-12.
54. Пиккарди Дж. – Химические основы медицинской климатологии. // Гидрометиздат. – Л. – 1967. – 96с.
55. Павлова Р.Н., Зайцева Н.К. – Сопоставление показаний унитиолового и акрилонитрилового тестов со спонтанными изменениями ферментативной активности нервной ткани. // Электромагнитные поля в биосфере. – Наука. – М. – 1984. – т.1. – с. 215-220.
56. Горшков Э.С., Шаповалов С.Н., Соколовский В.В., Трошичев О.А. – О гравитационной обусловленности флуктуаций скорости реакций окисления унитиола нитритным ионом. // Биофизика. – 2000. – т.45. – вып.4. – с. 631-635.
57. Горшков Э.С., Шаповалов С.Н., Соколовский В.В., Трошичев О.Н., Корнюшина Н.М. – Явление возникновения внешнеобусловленных регулярных флуктуаций скорости окислительно – восстановительных реакций. // Научные открытия. – Сб. кратких описаний науч. откр. – РАЕН. – М. – 2004. – с. 3-6.
58. Troshichev O.A., Gorshkov E.S., Shapovalov S.N., Sokolovskii V.V., Ivanov V.V., Vorobeitchikov V.M. – Variations of the gravitational field as a motive power for rhythmics of biochemical processes. // Adv. in Space Res., - 2004. – v.34. – issue 7. - p. 1619-1624.
59. Ivanov V.V., Sokolovsky V.V., Gorshkov E.S., Shapovalov S.N. and Troshichev O.A. – Periodical changes in the human urinary thiol content as related with cosmophysical factors. // Biophysics. – 2004. – v.49. – suppl. 1. – p. s 115-s 118.
60. Горшков Э.С., Трошичев О.А., Соколовская Т.М., Соколовский В.В. – О некоторых связанных с влиянием Солнца особенностях окисления тиолов в условиях высоких широт. // Слабые и сверхслабые поля и излучения в биологии и медицине. – 1-й Международный Конгресс. – Тез. докл. – СПб. – 1997. – с. 256-257.
61. Кириллов А.К. – Эффекты влияния ионосферы и ночной магнитосферы на космофизические флуктуации. // Междунар. Крымская Конф. «Космос и биосфера». – Тез. докл.– Крым.–2003.–с.212-213.