Адрес этой статьи в интернете: www.biophys.ru/archive/congress2012/proc-p173-d.htm
КОЛЕБАНИЯ БИОЛОГИЧЕСКОГО ВРЕМЕННОГО ПОЛЯ В ОРГАНИЗМЕ РАСТЕНИЯ
Наумов М.М.
Украина, Одесса, e-mail: naumovmm@te.net.ua
Краткий обзор основных теорий индивидуального развития растений. Данный обзор теорий составлен на основе работ Е.И. Ржановой [1], М.Х. Чайлахяна [2], В.С Цыбулько [3], Ф.М. Куперман [4], А.А. Шиголева [5,6], Д.А. Киризия [7], К.Дёрфлинга [8], Л.В. Белоусова [9], Б.Н. Белинцева [10], В.В. Полевого и Т.С. Саламатовой [11,12] и не претендует на всеобщность. Здесь, в таком обзоре, хотелось бы показать разнородную направленность теорий индивидуального развития растений.
А). В 1849 году В. Гофмейстер на основе анализа огромного фактического материала нарисовал отчетливую картину общего и различного в индивидуальном развитии растений, установил единство происхождения (гомологию) органов.
B). С появлением работ Ч. Дарвина начинают развиваться сравнительная морфология, анатомия и эмбриология. Индивидуальное развитие растений привязывают к истории вида. Онтогенетическое и филогенетическое развитие представляют двумя сторонами процесса развития. Филогенетическое направление впоследствии нашло свое выражение в биогенетическом законе.
C). Г. Клебс (1857-1918) исходил из представления о том, что весь ход развития растений, сроки наступления цветения и другие фазы развития связаны с внешними условиями. Ритм развития низших и высших растительных организмов зависит от факторов внешней среды, где доминирующую роль играют условия питания, свет (его интенсивность и качество), влажность почвы и воздуха, колебания температуры. Цветение наступает тогда, когда в тканях растений накапливаются сахара, и они преобладают над азотными соединениями. Г. Клебс пришел к заключению, что переход растения к плодоношению возможен только после того, как величина отношения C/N достигнет необходимого минимума.
D). Сакс, в 1880 году выдвинул гормональную теорию развития растений. Сакс предположил, что к моменту цветения в растениях происходит накопление особого цветообразующего вещества. Гипотеза Сакса о цветообразующих веществах не выдержала первой экспериментальной проверки.
E). Вент в 1920 году рассматривал развитие растений и отдельных органов на основании действия особых «органообразователей» (после открытия у растений гормонов – веществ высокой физиологической активности и работ по фотопериодизму): ризокалинов – корнеобразователей, каулокалинов – стеблеобразователей, филлокалинов – образователей листьев и так далее. Вент в своих опытах нашел, что при фототропизме ростовые гормоны смещаются в затененную сторону стебля, в результате чего происходит его положительный изгиб.
F). Н.Г. Холодный – 1928 год, рассматривал функцию регулятора на основе гормональной теории, обусловливающей морфогенез растений. Предпочтение отдавалось в основном одному гормону роста – ауксину. Была создана гормональная теория тропизмов, в основе которой лежат электрофизиологическая поляризация тканей вследствие одностороннего действия силы тяжести, света и других факторов. В результате, поляризация вызывает неравномерное распределение гормона роста и неодинаковую реакцию тканей стеблей и корней на действие этой поляризации.
G). Открытие фотопериодизма растений Гарнера и Алларда (1920-1923) показали, что процесс развития растений, переход к цветению зависит от периодичности смены дня и ночи. Растения в процессе эволюции приспособились не к свету вообще, а к его определенной периодичности. У одних растений увеличение длины дня ускоряет цветение и плодообразование, у других, наоборот, то же самое ускорение развития вызывает снижение светлого времени суток (укорочение длины дня). Различают растения короткого дня, длинного дня и нейтральные по отношению к светлому времени суток.
H). М.Х. Чайлахян в своих исследованиях (1937 год) использует гормональную теорию флоригена. На опыте показано, что возникают коррелятивные отношения между листьями и стеблевыми почками в течение развития растений. На основе этого, ищется материальный носитель, способный к транспорту по стеблю и направляющий формирование побегов в сторону репродуктивного развития. Делается вывод о гормональной природе процессов развития растений. Цветение растений и образование семян наступают в результате образования в листьях и передачи в почки достаточного количества гормона цветения. Как отмечает К. Дёрфлинг (1985 г.), наблюдаются безуспешные попытки выделить стимул цветения. В то же время, физиологи растений исследовали, как влияют на образование цветков фитогормоны и другие вещества. Большинство фитогормонов (ауксины, этилен, гиббереллины, абсцизовая кислота, цитокинины, стероиды) обладает широким спектром действия, но вопрос образования цветков еще не решен.
I). Исходя из основных возрастных этапов онтогенеза растений и считая, что развитие организма есть борьба и единство процессов старения и омоложения Н.П. Кренке (1940 г.) дал общий тип кривой изменчивости возрастных признаков в цикле жизни растений. Обоснованием является изменение морфологических признаков, таких, как величина и форма листовых пластинок и черешков, особенности строения ветвей, побегов и других органов. Изменениям, связанных с возрастом растений, изменениям морфологических признаков соответствуют закономерные физиологические и биохимические изменения.
J). В 1953 году Ф.М. Куперман выдвинула теорию этапов развития растений. Выделено 12 этапов органогенеза, при этом каждый этап характеризуется своей органообразовательной направленностью, зависимой от деятельности конуса нарастания (меристематической ткани) надземного побега растения. Каждый этап имеет свои, определенные требования к комплексу факторов внешней среды. На каждом этапе формируются определенные, морфологически тождественные органы, выполняющие одноименные функции в растительном организме. Изучение процессов формирования органов плодоношения злаков позволили выделить следующие основные этапы: I – формирование первичного конуса нарастания стебля; II – усиленная дифференциация зачаточных стеблевых узлов, междоузлий стебля и зачатков влагалищ стеблевых листьев; III – вытягивание конуса нарастания с одновременной дифференциацией сегментов в нижнем участке конуса нарастания и формированием кроющих чешуевидных листьев валиков; IV – формирование колосковых лопастей, выходящих их пазухи кроющих листовых чешуй; V – начало формирования колосковых и цветочных чешуй, а также зачатков пестика и тычинок; VI – формирование спорогенной ткани пыльцевых зерен и пестика; VII – растяжение в длину члеников колосового стержня и рост покровных органов колоска, цветков, а также остевидных образований или остей; VIII – выколашивание (выметывание); IX – цветение – оплодотворение; X – формирование зародыша и эндосперма; XI – восковая спелость; XII – созревание плода и семени злаков.
K). В агрометеорологии А.А. Шиголевым в 1951 году предложено цветение растений и другие фазы развития определять гипотезой о постоянстве сумм эффективных температур. Согласно этой гипотезе наступление фаз развития определяется необходимым количеством тепла. Гипотеза выражена уравнением:
|
|
где n – продолжительность межфазного периода; А – сумма эффективных температур (постоянная величина), представляющая собой сумму разностей между средней суточной температурой воздуха t и свойственным данному растению значением нижнего предела эффективной температуры В. За нижний предел для зерновых культур А.А. Шиголевым принята температура, равная 5 оС.
L). В 1976 году А.Л. Курсановым опубликован труд по теории донорно-акцепторных отношений в растении. Продукты фотосинтеза, образующиеся в фотосинтезирующих органах – донорах, перемещаются оттуда к потребляющим тканям – акцепторам. Такими тканями являются, в первую очередь, активные меристемы, включая камбий, а также ткани запасающих органов, которые часто образуют главную емкость для ассимилятов. Поток ассимилятов к потребляющим тканям ориентируется силой поляризующего фактора, который имеет тенденцию к затуханию по мере удаления от источника, посылающего сигнал. Природа аттрагирующего фактора до сих пор остается неясной. Поэтому распределение ассимилятов пока рассматривается в феноменологическом аспекте. Относительно природы «запроса» существуют три основные гипотезы. Первая их них исходит из того, что его носителями являются специальные вещества – метаболиты, выделяемые растущими клетками в тканях растения, например фитогормоны. Вторая гипотеза состоит в том, что скорость распространения электроимпульсов по проводящим пучкам больше соответствует скорости ответных реакций на «запрос» потребляющего органа, чем скорость передвижения фитогормона. Третья гипотеза заключается в том, что сила «запроса» определяется скоростью и полнотой использования тканью поступающих ассимилятов.
M). Л.В. Белоусов (1987) и Б.Н. Белинцев (1991) рассматривают процессы могфогенеза как самоорганизующуюся структуру. Рассматривается генетическая программа развития. В основе процесса самоорганизации лежит морфогенетическое поле. Как и для изучаемых в физике полях, для морфогенетического поля указывается его источник, генерирующий некоторое действие в своей окрестности. Характерным признаком, присущим морфогенетическим полям, является способность регулировать пропорции развивающейся структуры. Морфогенетическое поле связывается с генами.
N). Современная физиология растений (В.В. Полевой, Т.С. Саламатова – 1991 г.) рассматривает развитие растений как четыре возрастных этапа : 1. Эмбриональный – развитие зародыша от зиготы до созревания семени включительно. 2. Ювенильный (этап молодости) – включает в себя прорастание семян или органов вегетативного размножения и характеризуется накоплением вегетативной массы. Растение в этот период, как правило, не способно к половому размножению. 3. Зрелости и размножения – характеризуется готовностью к зацветанию, заложением репродуктивных органов, их ростом и развитием, формированием семян и плодов. 4. Старости и отмирания – период от полного прекращения плодоношения до естественной смерти организма.
Неоднозначность временных процессов в организме растений. Основной факт, о котором необходимо сказать, заключается в следующем. Длительность любого периода развития растений, в том числе всего онтогенеза, не остается постоянной величиной. Например, длительность вегетационного периода подсолнечника на юге Украины может колебаться от 80 до 120 суток. При этом здесь рассматривается календарная шкала времени, то есть физическое время. Если рассматривать начало вегетационного периода (посев) и конец вегетационного периода (полное созревание) как два события, происходящие в организме растения, то можно отметить следующее. События «посев» и «полное созревание» для организма растения в любых случаях являются одним и тем же. В этом смысле можно ввести биологическую ось времени, в которой продолжительность времени между двумя событиями, посев-полное созревание, будут иметь одну и ту же длительность времени. Так же как приведенные два события (вегетационный период) можно в этой шкале времени рассматривать межфазные периоды, этапы органогенеза или любые другие события, которые характеризуются одним и тем же биологическим явлением, происходящим в организме растения. Введение биологической оси времени ранее осуществлялось неоднократно. Прежде всего, это метод сумм эффективных температур и его различные модификации [5,6,13,14,15,16,17,18]. Введение биологической оси времени позволяет сделать вывод о существенном релятивизме временных процессов, проходящих в организме растений, когда время растягивается или сжимается по своей оси. Установлено, что изменение длительности онтогенеза или любых его отрезков определяется состоянием факторов внешней среды в каждый момент времени исследуемого периода [5,6,15,16,17,18]. В целом, состояние внешней среды, характеризуемое различными факторами, определяет скорость развития растений. Согласно нашим исследованиям [19,20] скорость газообмена СО2 определяет не только процессы роста, но и процессы развития. Следует отметить, что на скорость газообмена СО2 в целом влияет состояние внешней среды и возраст растений. Если рассматривать газообмен СО2 для данного возраста растения, то его скорость определится только лишь внешней средой. Поэтому из совокупности всех факторов внешней среды необходимо выбирать факторы, которые существенно влияют на газообмен. К таким факторам внешней среды относятся факторы света, тепла, влаги, минерального питания. Существует максимальная для данного вида (сорта) скорость газообмена СО2, которая считается наиболее оптимальной. При таком газообмене скорость роста растений максимальна. Но так же максимальна скорость развития. При снижении скорости газообмена СО2 по отношению к максимальной для данного возраста растений процессы развития будут замедляться. В связи с этим, существуют оптимальные условия внешней среды, существуют оптимальные значения факторов внешней среды для газообмена СО2. Таким образом скорость газообмена СО2 определяет биологическую ось времени.
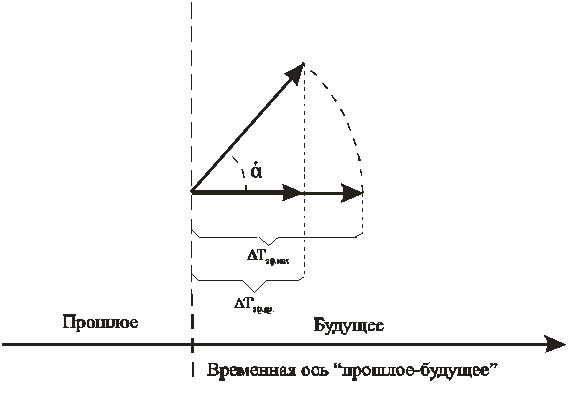
Двухмерность биологического времени. Здесь мы коснемся того, что существующие представления о биологическом времени характеризуют его двухмерность. Будем опираться на метод сумм эффективных температур, с успехом проверенный на практике[5,14,16,]. Рассмотрим элементарное приращение ΔТэф.отн. в методе сумм эффективных температур. Величина этого приращения зависит от температуры воздуха. Так, если температура оптимальна (20-220С для пшениц, 25-270С для подсолнечника, и т.д.), то приращение ΔТэф.отн. будет максимальным. Если температура упала до биологического нуля (50С – для пшениц, 8-10 0С для подсолнечника, и т. д.), то приращение ΔТэф.отн. будет нулевым за одни расчетные сутки. То есть мы имеем вектор, который в зависимости от температуры воздуха меняет свою длину от максимальной до нулевой при падении температуры от оптимальной до температуры нулевой биологической активности. Тогда мы имеем проекцию реального вектора ΔТэф.max, который в зависимости от температуры воздуха (напряженности фактора тепла) осуществляет поворот вокруг некоторой точки, рис. 1. Точка поворота вектора – это текущий момент времени. Угол α будет зависеть от температуры воздуха. При оптимальной температуре воздуха угол α будет равен 0 и, следовательно, приращение ΔТэф.пр. будет максимальным: ΔТэф.пр.= ΔТэф.мах.. При температуре воздуха равной биологическому нулю угол α будет равным π/2, и, следовательно, проекция на ось будет нулевая. Это даёт нулевое приращение ΔТэф.пр.: два вектора будут ортогональны. Из этого представления видно, что метод сумм эффективных температур рассчитывает сумму проекций реального вектора на физическую ось времени. В таком случае реальный вектор может поворачиваться как минимум в двухмерном временном пространстве, то есть во временной плоскости. В отличие от метода сумм эффективных температур можно рассматривать относительную ось биологического времени с теми же самыми выводами. При этом, можно рассматривать как фактор тепла так и другие факторы жизни либо их совокупное влияние.
|
|
|
Рисунок 1 – Представление элементарного приращения ΔТэф.отн. метода сумм эффективных температур в виде поворота реального вектора ΔТэф.max. ΔТэф.отн.=ΔТэф.пр.
|
Автономная система двух дифференциальных уравнений биологического временного поля растений. Разностная форма расчета оси биологического времени получена и проверена в работах [19,20]:
|
|
|
Тн≤Т≤Тцв, |
(1) |
|
|
Тцв≤Т≤Тсозр, |
(2) |
где Т – биологическая ось времени, изменяется за весь онтогенез от 0 до 2, отн. ед.;
ΔТopt – максимальное приращение биологической оси времени при оптимальных значениях факторов внешней среды, константа, отн. ед.;
Usr – уровень напряженности факторов внешней среды за время ΔТ, отн. ед., рассматривается как отношение газообмена всего растения при текущих условиях среды к газообмену растения в оптимальных условиях среды;
j – номер шага расчетного периода (номер суток).
Тн – начальное значение оси биологического времени, соответствующее биологическому времени, заключенному в семенах растений;
Тцв – точка оси биологического времени соответствующая половине онтогенеза, в наших исследованиях [19,20] соответствует моменту цветения;
Тсозр – момент завершения онтогенеза, соответствует фазе полного созревания.
Переход к дифференциальной форме выражения оси биологического времени можно осуществить в виде:
|
|
|
Тн≤Т≤Тцв, |
(3) |
|
|
Тцв≤Т≤Тсозр, |
(4) |
где dT – дифференциал биологической оси времени;
dTopt – дифференциал биологической оси времени при оптимальных значениях факторов внешней среды, показывает максимальную скорость развития;
dTвнешняясреда – дифференциал биологической оси времени в реальных условиях внешней среды;
Usr – уровень напряженности факторов внешней среды, отн. ед., показывает отношение газообмена всего растения при текущих условиях внешней среды к газообмену всего растения при оптимальных условиях внешней среды на данном шаге dT.
При таком переходе, от разностной формы расчетов к дифференциальной мы перешли от рассмотрения скорости суточного газообмена СО2 к изменению газообмена в течении суток, и вообще к изменению газообмена на бесконечно малых отрезках времени. Для расчета оси биологического времени по уравнениям (3) и (4) необходимо проинтегрировать эти уравнения:
|
|
Тн≤Т≤Тцв , |
(5) |
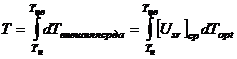 ,
,где ![]() -
среднее значение напряженности факторов внешней среды от момента времени Тн
до текущего значения Т.
-
среднее значение напряженности факторов внешней среды от момента времени Тн
до текущего значения Т.
Здесь мы проинтегрировали уравнение (3), при этом уровень напряженности факторов внешней среды Usr берется не для данного момента онтогенеза, а как некоторое среднее значение от начала момента интегрирования до текущего момента интегрирования. Тогда учтен весь комплекс факторов внешней среды, влияющих на скорость течения биологического времени на определенном отрезке интегрирования.
Среднее значение
уровня напряженности факторов внешней среды ![]() для
определенного отрезка интегрирования [Тн; Т] находится как
интеграл
для
определенного отрезка интегрирования [Тн; Т] находится как
интеграл
|
|
(6) |
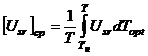 .
.Подставляя (6) в (5) мы получаем двойной интеграл со своей областью интегрирования
|
|
(7) |
где GF – квадратная область интегрирования со своей границей.
Здесь уже рассматривается пространство двух равноправных измерений биологического времени:
|
|
Интегрирование уравнения (3) естественным образом дало двухмерное пространство биологического временного поля.
Здесь следует отметить, что развитие растительного организма начинается с семени и заканчивается опять таки семенем, проходя все этапы, стадии и другие формы развития. В этом смысле нецелесообразно рассматривать пространство временных переменных TF и TR в декартовой плоскости. Структура биологического времени должна иметь замкнутый характер движений в связи с замкнутым процессом развития растений (от семени до семени) и повторением цикла онтогенеза при каждой вегетации. В связи с этим, в уравнении (7) мы перейдем к полярным координатам. То есть, мы будем рассматривать ту же самую декартову временную плоскость, только временные переменные будут выражены в полярных координатах:
|
|
(8) |
Область интегрирования GR будет представлять собой круг радиуса ρ. При этом полярные координаты имеют особую точку ρ=0. Такого значения ρ не может быть, так как в семенах растений всегда находится некоторое не нулевое, структурное образование биологического времени. В то же время, из области GR исключен радиус, лежащий в отрицательных значениях по оси х. В нашем случае ρ может меняться в пределах
|
|
(9) |
для отрезка биологического времени [Тн; 1].
Учитывая (9), интеграл (8) принимает вид:
|
|
(10) |
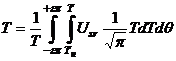 ,
,где θ – угол поворота вектора биологического времени;
ε – некоторое малое численное значение (отн. ед) показывающее в произведении επ содержание биологического времени в семенах растений, соответствует ρ0.
При переходе от уравнения (8) к уравнению (10) мы использовали следующую конструкцию: мы рассматриваем движения биологического времени в координатной плоскости переменных TF и TR; при этом считается, что физическое время имеет свойство движения равномерно слева направо; движения биологического времени осуществляются по кругу; точнее от -επ до +επ не совершая полного оборота; в этом случае взаимосвязь физического и биологического времен, как переход от (8) к (10) осуществится как интегрирование уравнения (10).
Теперь мы можем перейти непосредственно к построению автономной системы двух дифференциальных уравнений биологического временного поля. Рассмотрим подынтегральное выражение уравнения (10). Это дифференциальное уравнение двух переменных T и θ. Оно имеет вид определителя
|
|
(11) |
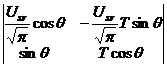
и, согласно работам Л.С. Понтрягина и А.А Андронова [21], являются вторыми производными дифференциальной системы уравнений:
|
|
(12) |
Нам остается найти явный вид системы дифференциальных уравнений (12). Для этого необходимо проинтегрировать один раз систему дифференциальных уравнений в частных производных (11) (см. [21]):
|
|
|
(13) |
Таким образом, мы нашли автономную систему двух дифференциальных уравнений в пространстве двух переменных: Т и θ. Здесь, в (13) переменная t является физическим временем, которое течет равномерно и прямолинейно. Система (13) определяет структуру биологического временного поля для первой половины онтогенеза. Для второй половины онтогенеза аналогично получим:
|
|
|
(14) |
Таким образом, мы получили две системы двух дифференциальных уравнений: (13) и (14) для двух отрезков онтогенеза до цветения и после цветения. Переход от одной системы уравнений (13) к другой системе уравнений (14) осуществляется в растении в момент цветения. Отметим так же и то, что полученные системы (13) и (14) согласно работе [21] являются грубыми динамическими системами. Грубые динамические системы заполняют области пространства динамических систем в целом. Грубые системы можно рассматривать как наиболее простые в соответствующем пространстве динамических систем [22]. Понятие грубости системы соответствует понятию надежности растительных систем [23].
Анализ полученных систем дифференциальных уравнений биологического временного поля. Теоретические результаты. Будем считать выполненными условия теорем существования грубой динамической системы биологического временного поля растений (13) [21]. Тогда мы получаем следующие теоретические результаты. В полной совокупности такая система дифференциальных уравнений биологического временного поля растений дает 11 топологических типов интегральных траекторий времени [21], табл. 1. Эта таблица получена на основе сопоставления результатов [21] и результатов цитологических исследований, изложенных в [24].
|
Таблица 1 - Совокупность интегральных траекторий системы биологического временного поля растений (13) в сопоставлении с клеточной структурой растений |
|||
|
I. Состояния равновесия: |
Узлы (фокусы) |
1 |
Рибосома |
|
Седла |
2 |
Хроматин |
|
|
II. Предельные циклы |
3 |
Клеточное ядро (управляющая структура) |
|
|
III. Сепаратрисы: |
Выходящие из узла (фокуса) или стремящиеся к узлу (фокусу) |
4 |
Митохондрия |
|
Сворачивающиеся с предельного цикла или стремящиеся к предельному циклу |
5 |
Хлоропласт |
|
|
Входящие в область GR |
6 |
Клеточная стенка |
|
|
IV. Траектории, имеющие в качестве своих предельных траекторий лишь узлы (фокусы) и предельные циклы, расположенные в области GR |
Выходящие из узла (фокуса) и стремящиеся к узлу (фокусу) |
7 |
Аппарат Гольджи |
|
Сворачивающиеся с предельного цикла и стремящиеся к предельному циклу |
8 |
Плазмодесма |
|
|
Выходящие из узла (фокуса) и стремящиеся к предельному циклу (или обратно) |
9 |
Лизосома |
|
|
V. Траектории, входящие в область GR и не являющиеся сепаратрисами: |
Стремящиеся к узлу (фокусу) |
10 |
Вакуоль |
|
Стремящиеся к предельному циклу |
11 |
Эндоплазматический ретикулум |
|
Кроме того, мы можем отметить, что динамическая система (13), а так же (14) разбивает область GR на конечное число связных ячеек – растительных клеток, заполненных обыкновенными временными траекториями. Такие клетки разбиваются на два класса: на класс клеток, примыкающих к циклу без контакта, и на класс внутренних клеток. Каждая такая клетка имеет в составе границы одну особую траекторию, являющуюся «элементом притяжения», или стоком, и одну особую траекторию, являющуюся «элементом отталкивания», или, источником. Каждая клетка имеет в составе границы одну особую траекторию, являющуюся стоком [21]. Будем считать, что для систем (13) и (14) существует взаимно однозначное и взаимно непрерывное преобразование области GR в себя. Тогда качественная структура разбиения этих временных областей на траектории определяется особыми траекториями, направлением движения по замкнутым траекториям и характером устойчивости предельных особых траекторий [21]. Это утверждение означает, что предельный цикл, отвечающий клеточному ядру, определяет управляющую структуру растительного организма. В то же время существуют и другие траектории, отвечающие другим растительным структурам (другим структурам клетки). К теоретическим результатам можно отнести так же и следующее. Временная структура по типу седла сопоставляется с хроматином клетки. Тогда, на основании [21], сепаратрисы (усы седла) в нормальном состоянии клетки не идут из седла в седло, рис. 2
|
|
||
|
а |
б |
в |
|
Рисунок 2 – Сепаратрисы седел. а – сепаратрисы идут из седла в седло, система (13) стает негрубой; б, в – сепаратрисы не идут из седла в седло, система (13) грубая (по данным [22]) |
||

Это означает, что клетка не делится и хроматин находится в свободном состоянии. Как только (с движением биологического времени) усы седла начинают идти из седла в седло происходит разрыв грубости системы - движение хроматина (образуется веретено), клетка делится, рис. 3.
Анализ полученных систем дифференциальных уравнений биологического временного поля. Практические результаты. Исследование системы уравнений (13) проводилось согласно методу, изложенному в работе [25]. Следует отметить, что системы (13) и (14) аналитически не интегрируются. Тривиальный случай: константы интегрирования с1, с2, с3 и с4 равные нулю не рассматриваются. Метод исследования системы (13) является методом конечных приращений для построения фазового портрета системы и соответствующих выводов. Фазовый портрет системы (13) для двух переменных Т и θ представлен на рис. 4.
Прежде всего, нужно
отметить, что система (13) разбивает область существования переменных Т и
θ на конечное число связных ячеек – растительных клеток. При этом в
полученных результатах присутствуют все 11 топологических типов интегральных
траекторий биологического временного поля. Интегральные траектории заполняют
всю область пространства GR. Интегральные траектории биологического временного поля
имеют всевозможные направления и пронизывают всю область определения, которая
соответствуют целостности растительного организма. Константы интегрирования с1
и с2 соответствуют культуре подсолнечника и составляют значения: с1=0.65
отн. ед. биологического времени и с2=0.28 отн. ед. биологического
времени. Эти константы могут быть записаны в виде: 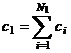 и
и
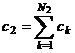 ,
где N1 и N2 числа, которые дают в сумме N1+N2 число генов растения, то есть
констант биологического времени.
,
где N1 и N2 числа, которые дают в сумме N1+N2 число генов растения, то есть
констант биологического времени.
|
|
|
Рисунок 3 – Митоз в клетках корешка алоэ (по данным [24]). |

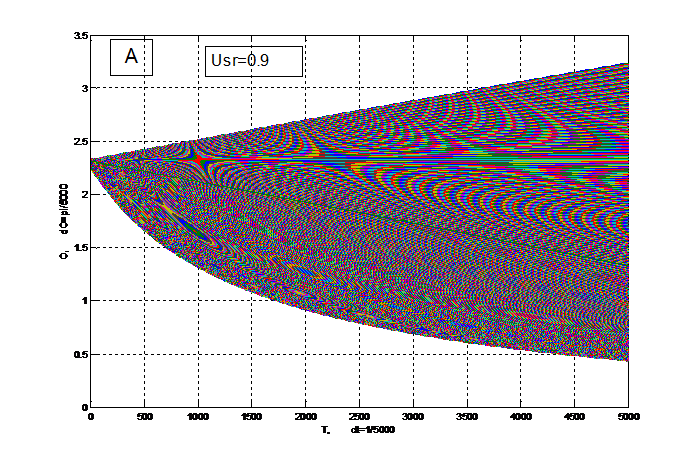
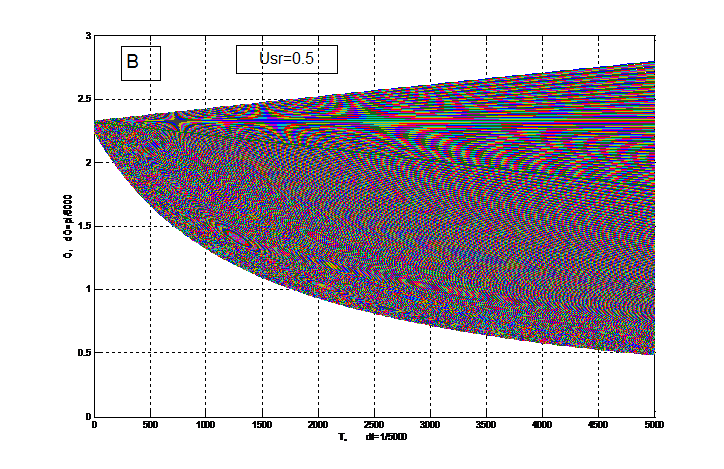
На рис. 4 вся область биологического временного поля разбита интегральной прямой на две подобласти: верхнюю и нижнюю. Верхняя подобласть соответствует надземной части растения, а нижняя подобласть соответствует корням. В верхней подобласти существует интегральная траектория (прямая) на которой последовательно расположены интегральные траектории по типу седла. При чем усы этих седел идут из седла в седло, что нарушает условия грубости динамической системы биологического временного поля на этих траекториях: происходит интенсивное деление клеток. Таким образом, эти приведенные траектории соответствуют меристематической ткани конуса нарастания надземного побега. Нулевые точки последовательных седел отвечают моментам начала заложения примордиев листьев и последнее седло отвечает заложению репродуктивного органа. Нижняя подобласть, соответствующая корням не содержит траекторий седел, где усы седла идут из седла в седло, что говорит об отсутствии в корнях управляющей структуры по типу конуса нарастания надземного побега. Вблизи нижней границы подобласти, соответствующей корням, на рис. 4 видно интенсивное образование мелких клеток – корневых окончаний.
В зависимости от состояния факторов внешней среды доля корней в целом организме растения меняется. Так при более напряженных условиях внешней среды, Usr=0.5 доля корней в организме растения увеличивается. В то же время снижается доля надземной части организма растения. Таким образом, уровень напряженности факторов внешней среды определяет рост и развитие организма растения.
|
|
|
|
|
Рисунок 4 – Фазовый портрет динамической системы биологического временного поля (13) для двух случаев: А – уровень напряженности факторов внешней среды Usr=0.9; В – уровень напряженности факторов внешней среды Usr=0.5. dt=Δt=1/5000 – шаг разрешения (15 мин), весь период времени от 0 до 5000 соответствует периоду времени 0.01≤Т≤1 или Тн≤Т≤Тцв (53-е суток); О=θ – угол поворота вектора времени. |
Заключение. Рассмотрение временных процессов в организме растения показывает, прежде всего, их релятивизм. Более того, направление временных процессов относительно оси физического времени крайне разнообразны: существует ровно 11 типов интегральных траекторий биологического времени и, на каждой траектории в силу их топологического различия будут проходить свои физиологические процессы. Существуют точки покоя биологического временного поля, источники и стоки биологического временного поля, определяющие направление физиологических процессов. В целом временное поле организма растения разбито на конечное число связных ячеек – растительных клеток и отвечает основным фактам роста и развития организма растения в зависимости от состояния факторов внешней среды, агрометеорологических факторов.
Список литературы
- Ржанова Е.И. Исторический обзор взглядов на онтогенез растений. Из-во МГУ, 1962, -56 С.
- Чайлахян М.Х. Регуляция цветения высших растений. М., «Наука», 1988, -559 С.
- Цыбулько В.С. Анализ экспериментальной обоснованности основных теорий и гипотез биологической природы фотопериодизма. // Физиология и биохимия культ. Растений, 1997, т. 29, № 4, С. 258-264.
- Куперман Ф.М., Дворянкин Ф.А., Ростовцева З.П., Ржанова Е.И. Этапы формирования органов плодоношения злаков. Из-во МГУ, 1955, -319 С.
- Руководство по агрометеорологическим прогнозам. Л., Гидрометеоиздат, 1984, т. 1, -309 С.
- Шиголев А.А. Руководство для составления фенологических прогнозов (озимая рожь, озимая пшеница, яровая пшеница, плодовые культуры, древесные растения лесных насаждений). М., Гидрометеоиздат, 1951.
- Киризий Д.А. Регуляция ассимиляции и распределения углерода в растении при изменении донорно-акцепторных отношений. // Физиология и биохимия культ. Растений, 1995, т. 27, № 4, С. 216-227.
- Дёрфлинг К. Гормоны растений. Системный подход. М., «Мир», 1985, -303 С.
- Белоусов Л.В. Биологический морфогенез. Из-во МГУ, 1987, -238 С.
- Белинцев Б.Н. Физические основы биологического формообразования. М., «Наука», 1991, -252 С.
- Полевой В.В. Физиология растений. М., «Высшая школа», 1989, -464 С.
- Полевой В.В., Саламатова Т.С. Физиология роста и развития растений. Л. Из-во ЛГУ, 1991, -239 С.
- Шнелле Ф. Фенология растений. Л., Гидрометеоиздат, 1961, -259 С.
- Шиголев А.А. Температура как количественный агрометеорологический показатель скорости развития растений и некоторых элементов их продуктивности. // Труды ЦИП, 1957, Вып. 53, С. 75-81.
- Бабушкин Л.Н. Оценка влияния погоды на скорость развития хлопчатника и других сельскохозяйственных культур и методы прогнозов наступления основных фаз развития их в условиях Узбекистана. Методические указания ЦИПа, 1951, Вып. 16, -47 С.
- Венцкевич Г.З. Из опыта работ по критической обработке фенологического материала агрометеорологических ежегодников. // Труды ЦИП, 1960, Вып. 98, С. 99-108.
- Дмитренко В.П. Использование влажности почвы и долготы дня в начале межфазных периодов для расчета их продолжительности. // Труды УкрНИГМИ, 1964, Вып. 44, С. 54-69.
- Дмитренко В.П. О совместном учете влияния факторов внешней среды на продолжительность межфазных периодов растений. // Труды УкрНИГМИ, 1987, Вып. 223, С. 3-23.
- Наумов М.М. Рост растений и биологическое время. // Вісник ОДЕКУ, 2005, Вип. 1, С. 72-78.
- Наумов М.М. Рост растений и биологическое время. Результаты расчетов. // Вісник ОДЕКУ, 2006, Вип. 2, С. 101-107.
- Понтрягин Л.С. Избранные труды. М., «Наука», 1988, т. 2, С. 30-34.
- Баутин Н.Н., Леонтович Е.А. Методы и приемы качественного исследования динамических систем на плоскости. М., «Наука», 1990, -486 С.
- Гродзинский Д.М. Надежность растительных систем. К., «Наукова думка», 1983, -367 С.
- Атабекова А.И., Устинова Е.И. Цитология растений. М., ВО «Агропромиздат», 1987, -246 С.
- Ризниченко Г.Ю. Лекции по математическим моделям в биологии. М-Ижевск, Изд. РХД, 2002, -236 С.
VIBRATIONS OF BIOLOGICAL TEMPORAL FIELD ARE IN ORGANISM OF PLANT
Naumov M.M.
Ukraine, Odessa, e-mail: naumovmm@te.net.ua
The brief review of basic theories of individual development of plants is given. An ambiguousness and two-dimensional of temporal processes passing in the organism of plant is shown. It is shown out and the off-line system of two differential equalizations of the biological temporal field is analysed. The theoretical and practical results of study of processes of the biological temporal field are given. The phase portrait of the system is built. The results of study give 11 types of integral trajectories of the biological temporal field, which are compared with 11 types of subcellular structures.