Адрес этой статьи в интернете: www.biophys.ru/archive/congress2012/proc-p111-d.htm
ДИСТАНТНЫЕ ВЗАИМОВЛИЯНИЯ БИОЛОГИЧЕСКИХ СИСТЕМ
Бурлаков А.Б., Бурлакова О.В., Голиченков В.А.
Московский государственный университет имени М.В.Ломоносова
Аннотация. Показано, что инкубация разновозрастных групп эмбрионов низших позвоночных в отдельных кварцевых кюветах при наличие только оптического контакта между группами приводит к изменениям параметров их дальнейшего развития по сравнению с контрольными группами. В зависимости от стадий развития взаимодействующих групп волновая биокоррекция может быть как положительной, так и отрицательной. Изменение спектральных характеристик и поляризация биоизлучений влияет на результат взаимодействия. Описаны различные аномалии развития, специфические для каждого сочетания взаимодействующих стадий и оптических условий коммуникации, вызванные дистантными волновыми взаимодействиями эмбрионов.
Ключевые слова: раннее развитие, костистые рыбы, бесхвостые амфибии, коррекция развития, дистантные взаимовлияния.
Одним из отражений химических энергетических процессов, происходящих в клетках, является эндогенное сверхслабое излучение, присущее всем биологическим системам. Еще в 1923 году А.Г.Гурвич сообщил об обнаруженном им очень слабом «ультрафиолетовом излучении, сопровождающем экзотермические химические реакции, идущие как в различных живых тканях, так и in vitro». По концепции А.Г. Гурвича и его последователей, излучение представляет собой специфический фактор, стимулирующий митозы, поэтому оно получило название митогенетического [1]. В работах А.Г. Гурвича и сотрудников впервые было показано дистантное, бесконтактное нехимическое влияние живых организмов друг на друга. В 1931 году Ж. и М. Магру [2,3] опубликовали данные об отрицательных эффектах дистантных воздействий бактериальных культур и молочнокислых ферментов на гаметы и на оплодотворенные яйцеклетки морских ежей, проявляющихся в угнетении последующего развития. После дистантного взаимодействия тех же бактериальных культур с эмбрионами комаров наблюдали, наоборот, ускорение вылупления личинок комаров [4]. Позже В.П. Казначеев с соавторами [5] показали, что при обработке цитотоксическими агентами (вирусами или сулемой) или при облучении ультрафиолетом одного из образцов культуры фибробластов в других образцах, инкубированных в отдельных емкостях так, что существовал только оптический контакт между образцами, наблюдали дегенеративные изменения клеток. Наличие дистантных взаимодействий продемонстрировано на дрожжевых культурах, на растительных тканях, на насекомых, на клетках крови человека [6] и у бактерий [7]. Химически и пространственно разделенные культуры микроорганизмов способны оказывать влияние на физиологическое состояние друг друга, приводя, в частности, к изменению таких параметров, как рост [8, 9, 10], как длительность lag-фазы, время существования одной генерации, плотность [11].
В последние десятилетия сверхслабые излучения разной длины волны от биологических объектов разного уровня организации - отдельных компонентов развивающегося куриного яйца [12], половых продуктов и зародышей рыб [13 ] были непосредственно зарегистированы с помощью фотоэлектронных умножителей (ФЭУ). Такое самопроизвольное излучение биообъектов - биофотонная эмиссия - по ряду признаков является когерентным, и может быть носителем информации при дистантных взаимодействиях биологических объектов
Роль биофотонного излучения в дистантном взаимодействии клеток и тканей продемонстрирована в опытах с эксплантатами лактирующих млечных желез мышей, в ходе которых проводили регистрацию спонтанного свечения в области 400-600 нм [14]. В этих опытах одни фрагменты железы (детекторы), реагировали изменением метаболической активности на химическую стимуляцию или ингибирование секреции белка других фрагментов ткани - индуктора, инкубируемых в отдельных кюветах, при существовании только оптического контакта между детекторами и индуктором. Параллельно возрастал уровень регистрируемой биофотонной эмиссии. Обмен волновой информацией между двумя изолированными культурами иммунных клеток-нейтрофилов через стенки кювет продемонстрирован в опытах российских и китайских исследователей [15,16], в которых проводилась с помощью двух ФЭУ синхронная регистрация биофотонного излучения отдельных образцов. Отмечали вспышку как спонтанной, так и люминол-зависимой люминесценции в одном из образцов при индуцировании в нем фагоцитарной активности добавлением определенных веществ, сопровождающейся "дыхательным взрывом" и генерацией активных форм кислорода. Эта вспышка инициировала те же явления в необработанной суспензии нейтрофилов, находившейся в оптическом контакте с "активированной" суспензией. При дистантных взаимодействиях культур микроорганизмов изменяются спектральные характеристики излучений на всех фазах существования колонии, в lag-фазу и фазу экспоненциального роста различия спектральных характеристик сверхслабых излучений контрольных и опытных колоний более выражены по сравнению с таковыми для стационарной фазы роста [10]. При дистантном (то есть в отсутствии химического и механического контакта) взаимодействии микробов наблюдаются изменения стимулирующего влияния красного и инфракрасного света на рост бактерий [11][. Показано, что информационным сигналом в подобных коммуникативных взаимодействиях может служить ультрафиолетовый [8, 9] и видимый свет [17, 10, 11]. В последние годы на эмбрионах вьюна была зарегистрирована биофотонная эмиссия как на разных стадиях развития [13, 18], так и при дистантном взаимовлиянии разновозрастных групп эмбрионов [19, 20].
I. ДИСТАНТНЫЕ ВЗАИМОВЛИЯНИЯ В РАННЕМ ОНТОГЕНЕЗЕ НИЗШИХ ПОЗВОНОЧНЫХ ЖИВОТНЫХ.
Нами уже в течение ряда лет проводится исследование дистантных волновых взаимовлияний ранних зародышей низших позвоночных животных [21-23]. Дистантное волновое взаимовлияние развивающихся зародышей низших позвоночных является весьма перспективной биологической моделью для исследования роли сверхслабых излучений живых систем. С одной стороны, здесь и источниками волновых сигналов, и детекторами такого воздействия выступают динамически усложняющиеся биологические системы. Усложнение включает появление многоклеточности в процессе дробления, сложных клеточных перемещений, дифференцировок и взаимодействий разных типов клеток с межклеточными структурами и между собой, что приводит к созданию отдельных органов, объединенных в разные функциональные системы организма. С другой стороны, - волновая информация, воспринимаемая организмом (детектором) и корригирующая начальные моменты формирования систем, в дальнейшем развитии реализуется как значительные их изменения. Последнее позволяет прослеживать участие волновых воздействий в процессах усложнения развивающейся системы и использовать их как инструмент в исследовании самих процессов самоорганизации. Нами была разработана методика регистрации биологического эффекта дистантных взаимовлияний эмбрионов низших позвоночных [21, 13]. Эксперименты проводили, помещая группы зародышей одной стадии в стандартные спектрофотометрические кварцевые кюветы 10х10х40 мм по 45-50 штук в опытах с
|
|
|
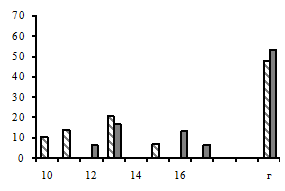
а б в г д |
|
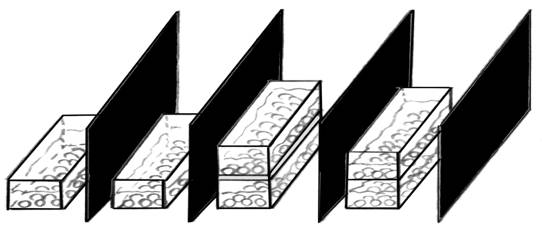
Рис. 1. Схема постановки эксперимента. а – кювета с контрольными зародышами «младшей» группы; б – кювета с контрольными зародышами «старшей» группы; в, г – подопытные пары кювет; д – непроницаемые для оптического контакта экраны |
вьюнами и по 25-30 штук, предварительно освобожденных от третичной оболочки, в опытах с амфибиями. Кюветы с экспериментальными и с контрольными зародышами соответствующих стадий размещалив большом металлическом боксе (стерилизаторе) для экранирования от внешних электромагнитных воздействий (рис.1).
Совместную экспозицию экспериментальных групп зародышей проводили в течение предварительно выявленного [13] оптимального времени – 18-24 часа. После этого переносили каждую группу зародышей в отдельные чашки Петри, затем максимально одновременно с помощью бинокулярной лупы МБС-9 определяли (1) распределение численности зародышей в соответствии со стадиями развития, (2) число погибших и (3) аномально развивающихся эмбрионов. Для определения стадий развития зародышей использовали таблицы нормального развития вьюна [24], травяной [25] , прудовой [26], шпорцевой [27] лягушек и серой жабы [28]. Статистическая обработка полученных данных проводилась при помощи компьютерной программы STADIA [51]. Для выявления достоверности различий между подопытными и контрольными группами зародышей использовались критерий Пирсона c2 и t–критерий, по парное сравнение оценивали по t–критерию Стьюдента.
1.1.Дистантные взаимовлияния у рыб
Были проведены исследования влияния оптических дистантных взаимодействий групп эмбрионов костистой рыбы вьюна (Misgurnus fossilis ).
Нами установлено, что содержание разновозрастных групп эмбрионов вьюна в отдельных кварцевых кюветах при наличии только оптического контакта между группами может приводить к изменениям параметров их дальнейшего развития по сравнению с контрольными группами. Такая биокоррекция развития может быть как положительной (ускорение и снижение числа аномально развивающихся зародышей по сравнению с контролем), так и отрицательной (замедление развития, увеличение числа аномальных зародышей, рост смертности) – рис.2.
К коррекции развития приводят дистантные взаимодействия только разновозрастных эмбрионов вероятно, излучение зародышей, не находя собственных воспринимающих систем, поглощается резонансными для этого излучения системами, имеющимися у зародышей другой стадии. Эффект дистантного взаимодействия эмбрионов определяется, прежде всего, сочетанием определенных стадий эмбрионального развития взаимодействующих групп особей - доноров и акцепторов излучения [21,22,13,29].
|
|
|
Рис. 2. Эффекты оптического контакта зародышей вьюна 0-й (15 мин после оплодотворения-«младшая» группа) и 1-й (2 бластомера-«старшая» группа) стадий развития. Положительная коррекция развития в «младшей» группе – ускорение, уменьшение числа аномальных (а) и погибших (г) зародышей. Отрицательная коррекция в «старшей» группе - замедление развития, увеличение числа аномальных и погибших зародышей в сравнении с контрольными группами. По оси абсцисс – стадии развития вьюна [24], по оси ординат – встречаемость в группе (%) |

Помимо общей коррекции развития результатом дистантного взаимовлияния может быть появление различных аномалий развития, специфических для каждого случая подбора взаимодействующих стадий. Особенно ярко это проявляется в нарушениях морфогенезов при
формировании головных и туловищных структур зародыша. Так, оптическое совмещение зародышей 0-1-й стадий с зародышами 8-й стадии (поздняя высокая бластула) вызывало появление зародышей с раздвоением головного конца (до 8% зародышей), причем уровень раздвоения мог быть в разной степени смещен к каудальной (хвостовой) области зародыша в зависимости от условий эксперимента. Такие аномальные зародыши впоследствии формировали двухголовых предличинок (рис.3) с развитыми головными структурами, с сердцами, работающими в одном или в различающихся ритмах. Следует отметить, что наши наблюдения за всю многолетнюю работу с ранним развитием вьюна показали, что “двухголовые” зародыши могут появляться в неблагоприятных условиях получения и инкубации икры (3-4 на 500-600 развивающихся эмбрионов). Но поскольку мы проводили эксперименты на икре высокого качества, подобные аномалии не встречались ни только в контрольных группах, но и во всех остальных партиях оплодотворенной и не вошедшей в эксперимент икры. Поэтому такая частота появления аномалий после определенных оптических контактов может считаться специфическим нарушением. После оптического контакта зародышей вьюна 5-й и 9-й стадий развития (рис.4) в «старших» опытных группах появлялись зародыши без хвостовых и с большим большим или меньшим недоразвитием туловищных структур (рис.5). Подчеркнем, что зародыши с возникшими после оптического контакта глубокими морфогенетическими нарушениями были достаточно жизнеспособны, проходили вылупление, жили в предличиночный период, т.е. такие значительные аномалии не вызывают катастрофических изменений и гибели всей системы – организм довольно долго живет и развивается. По-видимому, биоизлучение, избирательно нарушая начальные этапы механизма реализации самоорганизации определенной подсистемы (формирования осевых структурур зародыша), оказывается достаточно гармоничным для системы в целом.
|
А |
БА |
||||
|
В |
Г |
||||
|
Рис. 3. Предличинки вьюна: А – норма; Б, В и Г – раздвоение переднего отдела на разных уровнях после оптического контакта зародышей 0-1-й стадий с 8-й стадии. |
|||||




|
|
Б |
|
Рис. 4. Зародыши вьюна на 5 –й (А) и 9-й (Б) стадиях развития по таблицам нормального развития [24]. |
|


Удалось выявить, что характер проявления эффекта взаимного дистантного влияния связан с особенностями динамики развития зародышей вьюна. Для характеристики темпов развития зародышей каждой кладки мы проводили сопоставление достигнутых к определенному моменту стадий развития с теоретически рассчитанными по τо для данных температурных условий. Теоретическое значение стадии, соответствующей максимуму распределения зародышей по стадиям развития (моде распределения) в контрольных группах рассчитывали, используя безразмерную временную характеристику темпов развития низших развития зародышей каждой кладки мы проводили сопоставление достигнутых к определенному моменту стадий развития с теоретически рассчитанными по τо для
|
|
|
|
|
Рис. 5. Предличинки вьюна с недоразвитым туловищным и отсутствием хвостового отдела (А, Б) после оптического контакта. В – норма соответствующего возраста. |
||


данных температурных условий. Теоретическое значение стадии, соответствующей максимуму распределения зародышей по стадиям развития (моде распределения) в контрольных группах рассчитывали, пользуясь безразмерной временной характеристикой развития низших позвоночных τо [30]. Зная значение τо для температуры, при которой проводился эксперимент, и длительность эксперимента (T), рассчитывали величину T/τо,. По ее значению, пользуясь таблицами нормального развития, находили стадию, которой зародыши контрольной группы при нормальном развитии должны достигать ко времени окончания эксперимента. Эксперименты проводили на зародышах 0-й и 1-й стадий развития. Статистическая обработка результатов показала, что проведенные эксперименты можно разделить по характеру проявления эффекта на пять серий. Во взаимодействовавших группах одной серии влияния не проявились. В остальных обнаружили следующие эффекты: (1) положительную коррекцию развития “младшей” и отрицательную коррекцию развития “старшей” взаимодействовавших групп; (2) положительную коррекцию развития “младшей” группы и отсутствие различий подопытной и контрольной “старших” групп; (3) положительную коррекцию развития “младшей” группы и также положительную коррекцию развития “старшей”; (4) отсутствие различий подопытных и контрольных “младших” групп и отрицательную коррекцию развития “старшей” группы [31]. Выявлена четкая связь темпов развития зародышей и характера проявления эффекта их дистантного взаимовлияния. Группы, выделенные по характеру эффекта, оказались аналогичны группам опытов, разделенных в соответствии с темпами развития. При нарушении нормальных темпов развития вида, т.е. при отличии моды распределения по стадиям зародышей “младшей” и “старшей” контрольных (невзаимодействовавших) групп от рассчитанной стадии, достигаемой за время эксперимента при данной температуре, эффект дистантного взаимовлияния соответствующих экспериментальных групп зародышей проявлялся в меньшей степени, либо не проявлялся вовсе. Таким образом, эффект оптического взаимовлияния проявляется в случае соизмеримости темпов развития, т.е. продолжительностей последовательных событий эмбриогенеза. Чем продолжительней временной интервал (в составе общего времени дистантного взаимодействия), на протяжении которого взаимодействующие группы отвечают этому условию, тем сильнее выражается эффект взаимодействия. На степень проявления эффекта взаимодействия влияют темпы последовательных переходов зародышей от одного состояния к другому в ходе эмбриогенеза, характерные и индивидуальные для каждой из взаимодействующих групп зародышей. Эффект взаимодействия, вероятно, проявляется при достаточно длительной “компетентности” одной системы по отношению к излучению другой. Когда излучающая система развивается (и формирует определенный суммарный паттерн излучения) за время, отличное от времени развития (т.е. развертывания усложняющихся процессов) в воспринимающей системе, наблюдается нарушение или отсутствие эффекта дистантного взаимовлияния. В этом случае биоизлучение одних зародышей, вероятно, утрачивает определенное коррегирующее значение для других вследствие несовпадения по времени существования (развертывания) компетентных к этому излучению процессов (или динамических систем), что и выявили проведенные нами исследования [31,32].
Мы провели серию экспериментов по облучению зародышей вьюна разных стадий ИК-лазером (860 нм), интенсивность излучения которого модулировалась компъютерными записями спектров биофотонной эмиссии, предварительно зарегистрированной[1] с помощью ФЭУ в области 200-800 нм от зародышей вьюна в процессе раннего развития [33, 34, 35] для проверки временной специфичности характера биоизлучений и состояния воспринимающих излучения систем зародышей при оптическом контакте. Параметры лазерного излучения подбирали эмпирически таким образом, чтобы немодулированное излучение не оказывало действия на развивающихся зародышей. Длительность облучения - 10 минут - соответствовала истинному времени записи биофотонной эмиссии. Для модуляции интенсивности лазерного излучения использовали файлы эмиссии, зарегистрированные от неоплодотворенных яйцеклеток, через 15, 60, 70, 80 минут после оплодотворения, при появлении 2-х бластомеров и через 20 минут после образования 4-х бластомеров. Облучению подвергали вьюны трех стадий развития: неоплодотвореннаые яйцеклетки, яйцеклетки через 15 минут после осеменения (0 стадия развития) и стадия 2-4-х бластомеров. Контролем служили соответствующие группы зародышей, не подвергавшиеся облучению, и дополнительным контролем - соответствующие группы зародышей, облученных светом немодулированного лазера. Регистрацию биологического эффекта облучения в каждой группе проводили через 24, 48 и 65-70 часов после облучения. После облучения зародышей 0 стадии светом лазера, интенсивность которого модулировалась разными файлами, выявилось, что поражающим действием обладали файлы, зарегистрированные от зародышей на стадиях дробления (после образования 2 и 4-х бластомеров) - процент нормальных эмбрионов значительно снижался, что сохранялось до конца наблюдений. Модулирование лазерного излучения интенсивностью биофотонной эмиссии икры неоплодотворенной или оплодотворенной, но еще не дробящейся, не оказывало отрицательного воздействия на зародышей 0-й стадии. Однако, чувствительность зародышей к облучению одними и теми же файлами различалась в процессе их развития. Так, облучение светом лазера, интенсивность которого модулировалась файлом эмиссии, зарегистрированной от икры через 60 минут после оплодотворения, оказывало достоверное отрицательное действие на неоплодотворенную икру, но не вызывало достоверного ухудшения состояния зародышей при действии на зародышей 0-й стадий развития и 2-х и 4-хклеточных зародышей. Интересно отметить, что облучение икры через 15 минут после осеменения файлом эмиссии зародышей, находившихся на стадии 20 минут после образования 4-х бластомеров, вызывало большой процент уродств. Другие файлы не стимулировали таких резких отклонений от нормального развития. Зародыши на стадии 2-4-х бластомеров оказались мало чувствительными к облучению всеми файлами, по наблюдениям в первые двое суток развития. Однако, к моменту выклева в группах, облученных файлами 60 минут после оплодотворения и 2 бластомера, наблюдали достоверное снижение числа аномальных личинок. Проведенные эксперименты выявили, что модуляция лазерного излучения файлами биофотонной эмиссии зародышей разных стадий развития вызывала разный биологический эффект даже при использовании в качестве детектора зародышей одной и той же стадии развития. С другой стороны, облучение модулированным одними и теми же файлами лазером зародышей на разных стадиях развития также имело разные последствия. Таким образом, стадиоспецифичность реакции при оптическом взаимодействии зародышей разных стадий обусловлена как состоянием воспринимающей системы, так и спецификой биоизлучения донора.
1.2. Уровень свободнорадикальных процессов в зародышах вьюна при оптических контактах.
Характеристики излучения зародыша, как результат физико-химических процессов, протекающих в нем, зависят от его состояния, в том числе от интенсивности ростовых процессов. Свободнорадикальные процессы являются механизмом неспецифической активации многих клеточных функций, вплоть до клеточного деления, а также неспецифической регуляции транскрипции. Показано, что усиление радикалообразования в ядрах бластомеров бесхвостых амфибий предшествует удлинению клеточного цикла, а примерно десятикратный рост уровня свободных радикалов сопровождает активацию транскрипции в клетках [36]. Свободнорадикальный обмен, идущий в организме, клетке, имеет ритм, который чувствителен ко внешним резонансным воздействиям слабой интенсивности [37].
В ранние сроки экспериментов по оптическому контакту зародышей вьюна 0 и 1 (2 бластомера) стадий развития; 2 (8 бластомеров) и 28 (появление 13-й пары сомитов) стадий развития и 3 (32 бластомера) и 15 (обрастание бластодермой 2/3 поверхности желтка) стадий развития [24] проводили измерение интенсивности радикалообразования по уровню свободных радикалов (УСР) в клетках зародышей контрольных и подопытных групп методом привитой сополимеризации с использованием акриламида, меченного по углероду [ 38, 39]. УCH при контакте зародышей 0 и 1 стадии развития (рис.6) через 1ч 40 мин. не различался достоверно с соответствующими контролями в обеих взаимодействующих группах (181,69 и 156,93, соответственно, опыт и контроль в младших группах и 149,69 и 183,49 - в старших). В пробах, взятых через 3 часа 20 минут взаимодействия, УСР достоверно увеличивался в младшей группе по сравнению с соответствующими контрольными зародышыми (285,08 и 177,37, соответственно), тогда как в старших различия в опыте и контроле оставались недостоверными (268,16 и 218,37, соответственно). По окончании эксперимента эффект биокоррекции в этом опыте проявился как синхронизация и ускорение развития в младшей группе при снижении гибели с 36% до 28% по сравнению с
|
Младшая группа |
Старшая группа |
|
|
|
|
Рис. 6. Уровень свободных радикалов в зародышах вьюна в начальный период оптического контакта 0-й (младшая группа) и 1-й (старшая группа) стадий развития. По оси абсцисс – время опыта; по оси ординат – показатель УСР (распады в мин). |
|


контролем. В старших группах достоверных различий между показателями развития подопытной и контрольной групп не наблюдали. При взаимодействии зародышей стадии 2 (8 бластомеров) и стадии 28 (появление 13-й пары сомитов) достоверные различия уровня свободных радикалов в младшей группе по сравнению с контролем проявляются уже через 2 часа после начала оптического взаимодействия (1399 и 660, соответственно), и сохраняются через 4 часа (1536 и 667). В старших группах через 2 часа после начала опыта различия недостоверны (1584 в опыте и 1389 в контроле), но после 4 часов взаимодействия наблюдается достоверной снижение уровня свободных радикалов в зародышах опытной группы (2523 и 3568 в контрольной). После окончания эксперимента в младшей группе наблюдали синхронизацию и некоторое замедление развития по сравнению с контролем, а в старшей группе – значительную задержку развития (на 34,наиболее продвинутой, стадии находилось 86% зародышей в контрольной группе и только около 40% в опытной). При сочетании в эксперименте зародышей стадии 3 (32 бластомера) и стадии 15 (обрастание бластодермой 2/3 поверхности желтка) как через полтора, так и через 3 часа уровень свободных радикалов был достоверно выше в младших опытных группах (499,3 и 299,85) по сравнению с соответствующими контролями (352,6 и 177,3), а в старших наблюдали возрастание этого показателя в контроле (381,8 через 1, 5 часа и 459,05 через 3 часа) и снижение в опыте (299,15 и 223,85, соответственно). По окончании опыта в младшей опытной группе наблюдали ускорение развития, а в старшей – значительное замедление.
Таким образом, во всех экспериментах динамика уровня свободных радикалов в начале эксперимента коррелировала с конечным биологическим эффектом оптического взаимовлияния зародышей разных возрастных групп, что подтверждает роль свободнорадикальных реакций в эмбрионах как важнейшего эпигенетического фактора, определяющего готовность клеток к структурно-функциональным перестройкам и чувствительность к внешним воздействиям, в том числе и биоизлучениям.
1.3. Дистантные взаимовлияния развивающихся амфибий.
Были проведены исследования влияния оптических дистантных взаимодействий групп эмбрионов бесхвостых амфибий - травяной (Rana temporaria) и прудовой (R. esculenta) лягушек, серой жабы (Bufo bufo) и шпорцевой лягушки (Xenopus laevis), в период дробления, гаструляции и нейруляции. Прежде всего, подчеркиваем, что оптические контакты одновозрастных зародышей не вызывали достоверных изменений параметров дальнейшего развития по сравнению с развитием соответствующих контрольных групп, как это было установлено нами ранее и на зародышах вьюна. Результаты экспериментов, выполненных на зародышах травяной лягушки, показали, что чаще всего наблюдается отставание зародышей «младшей» группы (замедление развития) от зародышей контрольных групп и/или опережение зародышей «старшей» из контактировавших групп зародышей соответствующего контроля в среднем на 1-2 стадии (рис.7). Однако, сочетания определенных стадий, например, младшая группа 5-6 стадии (16-32-х-клеточные зародыши) и старшая – от 10 стадии, непосредственно предшествующей гаструляции, и до стадии средней гаструлы (13 стадия); или совмещение еще не дробящихся зародышей (0-1 стадии) с 8-миклеточными (4 стадия развития) вызывало достоверное ускорение развития зародышей и младшей группы. Во всех экспериметальных группах зародышей травяной лягушки число аномальных и погибших зародышей не меняется достоверно по сравнению с с контрольными группами, однако единичные проявления некоторых аномалий развития были характерны только для зародышей подопытных групп после прямого оптического контакта.
На наиболее ранних стадиях
(дробящиеся яйцеклетки вплоть до бластулы) замедлялось развитие, но при этом
достоверно снижался процент аномальных зародышей после взаимодействия с
зародышами на стадиях гаструляции, которые реагировали ускорением развития
(рис.8,А). Причем такое ускорение часто сопровождалось увеличением числа
аномальных зародышей. После оптических контактов таких ранних зародышей с более
старшими зародышами (на стадиях поздней нейрулы - хвостовой почки) младшие
ускоряли развитие, а старшие – замедляли (рис.8,Б). К числу характерных аномалий,
появляющихся в экспериментальных группах, можно отнести устойчивые искривления
туловища за счет асимметрии развития левой и правой частей на разных уровнях
переднезадней оси зародыша (рис.9Д, Е), нарушение развития головных структур
(рис.9Е). Среди аномальных особей экспериментальных групп наиболее часто
встречаются отеки вентральных отделов – аномалии, наблюдающиеся и в контрольных
группах, но значительно реже (до 2-4% в контроле и до 20% - после оптических
контактов на стадиях поздней бластулы – средней гаструлы). Наибольшую
вариабельность стадиоспецифических ответов на оптические контакты выявили
эксперименты на зародышах серой жабы. Однако особенностью реакции этого вида
было увеличение
числа аномальных зародышей после оптических контактов во всех экспериментальных
группах (как на фоне любых проявлений коррекции и в «младших», и в «старших»
контактирующих группах) по сравнению с соответствующими контрольными (рис.10).
В связи с этим интересно, что этот вид имеет характерную форму кладок икры,
которые представляют собой «цепочку» икринок в студенистой третичной оболочке
(галерте), развешенной длинной нитью на подводной растительности. В результате
такого взаимного расположения, в норме зародыши жабы имеют минимальный
оптический контакт друг с другом, по сравнению с кладками лягушек. Не является
ли такая форма кладки эволюционным приспособлением для уменьшения площади
оптического контакта?
|
Младшая группа |
Старшая группа |
||
|
|
ан г |
||
|
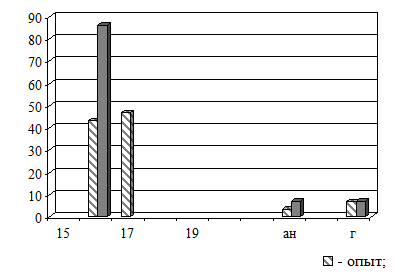
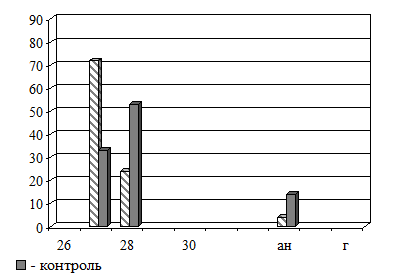
Рис.7. Эффекты оптического контакта зародышей травяной лягушки 3-й (4 бластомера) и 10-й (поздняя бластула) стадий развития. По оси абсцисс – стадии развития [25], ан – аномальные и г – погибшие зародыши; по оси ординат – встречаемость в группе (%) |
|||
|
Младшая группа |
Старшая группа |
||
|
|
|
||
|
|
|
||
|
Рис. 8. Эффекты оптического контакта разновозрастных групп зародышей прудовой лягушки: А – 7-й (морула) и 12-й (ранняя гаструла) стадий развития, Б – 7-й и 21,5-й (поздняя нейрула) стадий развития. По оси абсцисс – стадии развития [26], ан – аномальные и г – погибшие зародыши; по оси ординат – встречаемость в группе (%). |
|||




|
Rana temporaria |
|
|
|
|
|
Rana esculenta |
|
|
|
|
|
|
|
|
Рис. 9. Основные типы аномалий развития зародышей травяной (Б) и прудовой (Г-Е) лягушек после оптических взаимовоздействий. (А и В – нормальные личинки). |
|






Нам впервые удалось проследить изменения динамики развития и определенные нарушения морфогенеза после длительных оптических контактов разновозрастных ранних зародышей шпорцевой лягушки – вида, наиболее используемого ныне как объект лабораторных исследований в биологии развития. Предварительные исследования показали, что экспериментальные воздействия не только оказывали влияние на изменение темпов последующего развития, но и вызывали появление аномалий (рис. 11) – микрофтальмию (А), одностороннюю анофтальмию (Б), раздвоение осевых структур и формирование личинки с раздвоенным хвостом (В). Т.е. спектр аномалий у этого вида, как и у вьюна, гораздо шире, чем у ранее исследованных амфибий.
В целом, воспроизводимость результатов по влиянию оптических контактов разновозрастных зародышей амфибий на их дальнейшее развитие высока и специфична для определенных сочетаний стадий развития.
1.4.Межклассовые оптические контакты и их последствия.
Исследовали последствия оптического контакта зародышей вьюна с зародышами бесхвостых амфибий (серой жабы, травяной, прудовой и шпорцевой лягушек). В целом, коррекция развития при всех сочетаниях видов наблюдалась в обеих взаимодействовавших группах, причем при всех межклассовых сочетаниях отмечалась сходная зависимость от возраста экспериментальных животных. Оптические контакты оплодотворенных и дробящихся яйцеклеток имели положительный коррегирующий эффект, тогда как зародыши, включенные в эксперимент на стадиях средней бластулы и старше, отставали в развитии от контроля. И, наконец, зародыши поздних стадий (поздняя нейрула и старше) при оптическом контакте с ранними стадиями (оплодотворенные или дробящиеся яйцеклетки) опять несколько ускоряли их развитие. Например, сочетание в опыте травяной лягушки и вьюна,
находящихся на стадиях гаструляции, приводило к отрицательной коррекции развития обеих экспериментальных групп по сравнению с контрольными (рис.12).
|
А
|
Б
|
|
В
|
|
|
Рис. 11. Аномалии развития шпорцевой лягушки в группах после оптических взаимовлияний: микрофтальмия (А), односторонняя анофтальмия (Б), раздвоение хвостового отдела (В). Г- нормальная личинка того же возраста. |
|
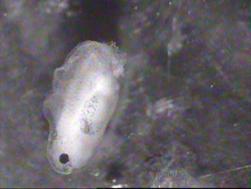



Оптический контакт между оплодотворенными яйцеклетками вьюна и шпорцевой лягушки (4 - 8 бластомеров) приводил к некоторому ускорению развития в обеих группах (рис.13,А), тогда как между оплодотворенными яйцеклетками вьюна и шпорцевой лягушкой на стадии бластулы (8 стадия) – к некоторому ускорению развития вьюна и отставанию в развитии зародышей лягушки (рис.13,Б).
После эксперимента с оплодотворенными яйцеклетками вьюна и прудовой лягушкой на стадии ранней гаструлы (рис.14,А) наблюдали некоторое отставание зародышей вьюна от контрольных групп, тогда как сочетание вьюна на стадии 4-х – 8-ми бластомеров с прудовой лягушкой на стадии хвостовой почки несколько ускоряло развитие вьюна (рис.14,Б).
|
R.temporaria |
M.fossilis |
|
|
|
|
|
|


При оптическом контакте зародышей вьюна и серой жабы отметили ранее не встречавшиеся аномалии в развитии. В этой серии экспериментов кварцевые кюветы с зародышами вьюна 14-й стадии развития (средина обрастания бластодермой поверхности желтка) совмещали с кюветами с зародышами жабы, находившимися на заключительных стадиях эмбрионального развития (перед выходом из зародышевых оболочек). Через 3 суток после окончания эксперимента у жаб наблюдали достоверное отставание в развитии, появление
|
|
|
А |
|
|
|
Б |
|
Рис.13. Результат оптического контакта шпорцевой лягушки и вьюна, находящихся на 3-й и 0-й стадиях, соответственно (А), и на 8-й и 0-й стадиях (Б) развития. По оси абсцисс – стадии развития (для лягушки [27], для вьюна [24]), г – погибшие зародыши; по оси ординат – встречаемость в группе (%). |
||



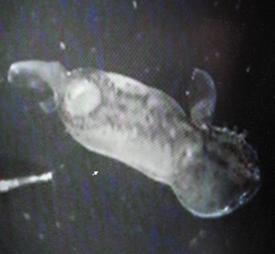
аномальных зародышей (в основном, отмечали нарушения формирования осевого комплекса), примерно 5% личинок погибли. Зародыши вьюна в опытных группах несколько опережали контрольных в развитии, но почти все (80.5%) были аномальными. Такие отклонения в развитии прежде не отмечались нами у развивающихся вьюнов – личинки были заметно крупнее контрольных личинок (длина тела личинки экспериментальных групп
4.94 + 0.14 мм; контрольных групп - 4.14 + 0.19мм), практически лишены пигментации глаз и пигментных клеток – меланофоров, многочисленных на этой стадии в норме, и были малоподвижными. У 15.2% личинок вьюна в экспериментальных группах проявилось искривление осевого комплекса. Аномалии развития не регулировались, личинки еще через сутки погибали.
Характер проявления эффектов оптического контакта зародышей амфибий отличается от такового для зародышей вьюна. Возможно, это связано с тем, что сами рыбы и амфибии различаются по таким признакам как соотношение желтка и свободной цитоплазмы в яйцеклетках (полилецитальные яйцеклетки у рыб - мезолецитальные у амфибий), тип дробления (дискоидальное у первых и полное неравномерное – у вторых). Все это создает разные условия для сверхслабых излучений этих объктов во внешнюю среду. Что касается их чувствительности к внешним биоизлучениям, то можно предположить, что различия вьюнов и амфибий по пигментации, которая у вьюнов обусловлена каротиноидами, а у амфибий – меланином, также накладывает отпечаток на проявление биокоррекции при оптических контактах эмбрионов. Так, пигмент меланин обладает свободнорадикальными свойствами, он является и универсальной «ловушкой» для фотонов, и протектором, связывающим избыток свободных радикалов. Молекула меланина не только накапливает, но и отдает их при молекулярных конформациях. Возможно, при этом спектральные и амплитудные характеристики излучения зародышей в чем-то нивелируют стадийную специфичность, которая обусловлена усложнением обменных процессов развивающегося зародыша. Некоторые основания к таким умозаключениям дают сопоставления специфики биологических эффектов оптических контактов зародышей разных таксономических групп. В этом ряду вьюны – наиболее лабильный вид, а среди амфибий – шпорцевые лягушки (наименее пигментированные). Разнообразие и стадиоспецифичность реакций более выражены у прудовой лягушки, чем у более пигментированной травяной и наиболее богатой меланином жабы. Возможно, особенности размножения этих видов сформировались и с учетом оптической коммуникации икры и зародышей в кладках как одного из факторов эволюции.
|
|
|
А |
|
|
|
Б |
|
Рис.14. Результат оптического контакта прудовой лягушки и вьюна, находящихся на 11-й и 0-й стадиях, соответственно (А), и на 3-й и 0-й (Б) стадиях развития. По оси абсцисс – стадии развития (для лягушки [26], для вьюна[24]), , г – погибшие зародыши; по оси ординат – встречаемость в группе (%). |
||




II. ВЛИЯНИЕ ИЗМЕНЕНИЯ ОПТИЧЕСКИХ ХАРАКТЕРИСТИК ИЗЛУЧЕНИЯ РАЗВИВАЮЩИХСЯ ЗАРОДЫШЕЙ НИЗШИХ ПОЗВОНОЧНЫХ НА РЕЗУЛЬТАТ ИХ ОПТИЧЕСКОГО КОНТАКТА.
2.1.Поляризация биоизлучения
Оптическая активность биологических макромолекул дает основание исследовать вопрос о поляризационной составляющей информации, передаваемой при дистантной биокоммуникации организмов. Вычленение поляризационной составляющей биооптического
излучения путем разделения взаимодействующих объектов поляризационными мембранами показало ее важную роль в изменении характера развития зародышей. При разделении кварцевых кювет с зародышами вьюна поляризатором (в видимом диапазоне) мы наблюдали значительные изменения эффекта дистантного взаимодействия [40-42]. Эксперименты проводили по обычной схеме, использовали два параллельных контроля: (1) группы зародышей соответствующих стадий в изолированных друг от друга кюветах и (2) группы, взаимодействующие без поляризатора. При оптическом контакте зародышей вьюна 6-й (бластула) и 0-й стадий развития в старшей экспериментальной группе как прямое взаимодействие с младшей, так и через поляризатор, не выявило достоверных отличий от не взаимодействовавшего контроля; в младшей - без поляризатора наблюдали значительное достоверное (р < 0,01) отставание в развитии и увеличение процента гибели зародышей по сравнению с соответствующим контролем. При разделении кювет со взаимодействующими группами поляризационной пластиной степень отрицательной биокоррекции развития зародышей младшей группы достоверно снижалась (р<0,01), но во всех случаях зародыши экспериментальных групп значительно отставали от невзаимодействующего контроля (р<0,001). Наименьший отрицательный эффект взаимодействия наблюдался в группах, разделенных поляризационной пластиной, вектор которой был расположен параллельно продольной оси кюветы. Минимальное отличие от групп, взаимодействовавших без поляризатора, наблюдали в экспериментальных группах при расположении вектора поляризатора перпендикулярно к продольной оси кювет. При смещении поляризатора под 45° биологический эффект по скорости эмбрионального развития был промежуточным (между 0° и 90°). Необходимо отметить, что левое и правое смещение вектора поляризатора относительно продольной оси кювет не давало идентичных результатов. Таким образом, результаты экспериментов по влиянию одного поляризатора показали, что степень проявления эффекта коррелирует прежде всего, с ориентацией поляризатора относительно продольной оси кюветы.
При использовании двух поляризаторов (поляризатора и анализатора) [13] в экспериментах на зародышах вьюна тех же стадий развития (0-я и 6-я) разделение кювет параллельно ориентированными по вектору поляризатором и анализатором приводило к некоторому улучшению состояния зародышей младшей возрастной группы по сравнению с прямым взаимодействием (р < 0,01), хотя процент погибших эмбрионов был выше. В старшей возрастной группе темпы развития отличались от не взаимодействовавшего контроля в меньшей степени, чем от взаимодействовавших напрямую зародышей. Однако в этой группе выявилось большое число (до 15-20% в каждом отдельном эксперименте) аномально развивающихся зародышей. Аномалии касались, в основном, формирования осевых структур. Это выглядело как нарушение замыкания бластопора, а при дальнейших наблюдениях такие отклонения приводили к искривлению и локальному раздвоению осевого комплекса. Разделение кювет с взаимодействующими группами скрещенными под 90 градусов поляризаторами в младшей группе почти полностью нивелировало эффект дистантного взаимодействия (недостоверное отличие от невзаимодействовавшего контроля) по скорости развития, хотя число погибших зародышей было достоверно выше, чем в контроле (р<0,05). В этой группе в каждом эксперименте регистрировали до 5-7% аномальных зародышей, погибавших в течение следующих суток. В старшей группе в этой серии опытов выявили рассинхронизацию развития, но, в целом, ситуация по темпам развития промежуточная между невзаимодействовавшими группами и взаимодействовавшими без поляризаторов. Аномалий развития не выявили.
Эксперименты на зародышах прудовой лягушки [42] на стадиях средней и поздней гаструлы (ст.13 и ст. 18 [26], соответственно) показали, что в группах, взаимодействовавших без поляризаторов наблюдалась биокоррекция развития: в младшей возрастной группе положительная, выражающаяся в некотором ускорении и синхронизации развития, а в старшей - отрицательная, выражающаяся в замедлении и рассинхронизации развития зародышей в группе по сравнению с не взаимодействовавшими контрольными группами. Но в младшей группе после взаимодействия выявились некоторые аномалии в развитии, касающиеся, в основном, структур осевого комплекса. Поляризаторы между взаимодействующими группами в старших возрастных группах незначительно улучшали ситуацию по скорости и синхронности развития по сравнению с группами, взаимодействовавшими без поляризаторов, но при взаимно перпендикулярном положении поляризаторов появлялись аномально развивающиеся и погибшие зародыши. В младших возрастных группах взаимодействие через поляризаторы усиливает отрицательную биокоррекцию развития, причем параллельное взаиморасположение векторов поляризаторов более эффективно, чем перпендикулярное. До 10% зародышей погибает при использовании поляризаторов. Таким образом, поляризация биоизлучения приводит к изменению биологического эффекта взаимодействия, причем степень таких изменений зависит от ориентации поляризаторов. После взаимодействия через поляризаторы у зародышей может проявляется ряд специфических аномалий морфогенеза, причем локализация и степень проявления некоторых нарушений коррелирует со степенью поляризации биооптического излучения.
Когда кюветы с экспериментальными разновозрастными группами зародышей вьюна были разделены одним ИК-поляризатором [41], т. е. оптический контакт между группами осуществлялся в ИК–диапазоне поляризованным излучением, результаты эксперимента значительно отличались от описанных выше даже при оптическом контакте тех же стадий.
При экспериментах с зародышами вьюна 0-й и 6-й стадий в младшей группе после оптического контакта через ИК-поляризатор произошла практически полная синхронизация (рис.15), но при этом заметно возросла гибель зародышей. В старшей группе наблюдается так же, как при поляризации в видимом диапазоне, но в более значительной степени, четкая бимодальность распределения зародышей по стадиям развития. Интересно, что коррекция в направлении формирования такой бимодальности под действием оптически модифицированных биоизлучений возможна на стадии бластулы, когда собственно, начинает проявляться морфогенетическая функция ядер нового организма.
|
Младшая группа |
Старшая группа |
|
|
|
|
А |
|
|
|
Б |
|
Рис.15. Влияние ИК-поляризаторов на проявление результата оптического контакта зародышей вьюна, находящихся на 0-й и 6-й стадиях, соответственно (А), и на 0-й и 14-й (Б) стадиях развития. По оси абсцисс – стадии развития [24], г – погибшие зародыши; по оси ординат – встречаемость в группе (%). |
||




Эксперименты с ИК-поляризацией биоизлучений подтвердили стадиоспецифичность эффектов оптических взаимовлияний. Следует подчеркнуть, что эффекты ИК-поляризатора не связаны с перекрыванием воздействия, поскольку состояние зародышей экспериментальных групп отличается от невзаимодействующего контроля.
2.2. Влияние изменений спектральных характеристик биоизлучений на результат оптических взаимовлияний развивающихся зародышей низших позвоночных.
В результате многочисленных экспериментов с использованием светофильтров различных типов было установлено, что дистантное взаимодействие эмбрионов возможно в различных областях как видимой части спектра, так и в ультрафиолетовом и ИК-диапазонах [42]. Было показано, что разделение разных групп эмбрионов светофильтрами определенным образом влияет на их эмбриональное развитие. Так, почти все широкополосные светофильтры серии БС ( БС-12, БС-8, БС-7, БС-4, БС-3, отрезающие более коротковолновую часть ультрафиолета) особенно не влияют на выживаемость эмбрионов вьюна на стадии ранней бластулы при их оптическом контакте с вьюнами, находящимися в начале развития (0-я - 4–я стадии), одновременно снижая гибель эмбрионов в младшей группе. Но в последней группе во всех случаях наблюдается четкое достоверное замедление скорости эмбрионального развития по сравнению с соответствующими контрольными группами. При оптическом контакте вьюнов тех же стадий через желтые светофильтры (ЖС-16, ЖС-18) достоверно ( p<0,01) увеличивается почти в 3 раза гибель эмбрионов в группе, исходящей от ранней бластулы, и практически не меняется при этом смертность в группе, исходящей от начала развития.
Для более четкого анализа спектральных характеристик биоизлучения [43] использовали интерференционные светофильтры с шириной полосы пропускания ±3 нм и максимумами пропускания при 372, 379, 386, 401, 406, 416, 455, 482, 526, 546, 593, 628, 756 и 780 нм. Во всех случаях контрольные группы, размещенные как на, так и под светофильтрами и совмещенные оптически с кюветой без живых объектов, развивались впоследствии, как и в отдельно стоящих контрольных кюветах, т.е. сами светофильтры как со стороны зеркальной, так и с противоположной достоверно не оказывали влияния на параметры развития зародышей. Результаты проведенных экспериментов позволили выявить два интересных момента. Во-первых, использование интерференционных светофильтров, вычленяющих разные длины волн, при оптическом контакте одних и тех же возрастных групп эмбрионов имеет разные биокоррегирующие последствия. Т.о. биоизлучение проявляет себя в разных областях оптического спектра. Во-вторых, сочетание разных возрастных групп зародышей дает разные результаты при использовании одного и того же светофильтра. Эти факты могут еще раз свидетельствовать, с одной стороны, о неодинаковом характере сверхслабого излучения, продуцируемого эмбрионами разных стадий развития, а с другой, - о возможных возрастных изменениях воспринимающих эти излучения систем. Так, например, зародыши, помещенные в эксперимент на 6 стадии развития на излучение одной и той же длины волны, проходящее через светофильтр 593 нм от зародышей 0 стадии, реагируют положительной коррекцией развития, а от зародышей 30-31 стадий – слабым угнетением. Дополнительную серию экспериментов провели на зародышах ранних стадий вплоть до поздней (эпителиальной) бластулы, оценивая последствия оптических контактов через интерференционные светофильтры групп зародышей, находящихся на одной стадии [44]. Следует отметить, что все наши предыдущие эксперименты свидетельствовали о том, что прямые оптические контакты одностадийных зародышей достоверно не изменяли параметры последующего их развития, т.к. и характер биоизлучений, и состояние воспринимающих систем стадиоспецифичны, т.е. в достаточной степени сходны у зародышей одной стадии (в пределах индивидуального и внутристадийного разброса). Однако, проведенные эксперименты по оптическим контактам групп одностадийных (одновозрастных) зародышей, находящихся на самых ранних стадиях развития (стадия 1- двух клеточный зародыш; стадия 3 и 3,5 – соответственно, 32-х и 64-х клеточный зародыш и стадии 6), показала, что вычленение определенной длины волны негативно сказывается на выживаемости и степени синхронизации развития зародышей в группе. В группах одновозрастных эмбрионов вьюна, помещенных в эксперимент на начальных стадиях дробления и разделенных интерференционными светофильтрами с полосами пропускания 401нм, 482 нм или 546 нм, по окончании эксперимента до 8% зародышей развивались со значительными нарушениями морфогенеза, что в дальнейшем приводило к появлению специфических аномалий развития (рис.16), а при использовании светофильтров 482 нм и 546 нм еще и значительно увеличивалась смертность в группах (до 20%).Эмбрионы, помещенные в экспериментальные условия на 8 стадии развития, оказались чувствительными к пропусканию излучения через светофильтры 372 нм и 379 нм – мы наблюдали значительное увеличение смертности (до 30% в некоторых группах) по сравнению с контрольными. Оптический контакт через светофильтр с пропусканием в области 546 нм тормозил развитие, тогда как в 406 нм, напротив, способствовал слабому ускорению. Более выраженный эффект положительной коррекции с использованием светофильтров с максимумами пропускания не только в области 406, но и 401 нм мы наблюдали в
|
А |
Б |
В |
|
|
|
|
|
Рис.16. Типичные аномалии развития в группах одновозрастных вьюнов, помещенных в эксперимент на начальных стадиях развития, разделенных ИС с полосами пропускания 401нм, 482 нм или 546 нм. А - образование 2-х неравноценных голов и недоразвитие туловищных и хвостовых отделов; Б - недоразвитие туловищного и хвостового отдела; В – норма. |
||



экспериментах с эмбрионами 9 стадии развития (предшествующая процессу гаструляции стадия поздней эпителиальной бластулы) – явное ускорение на фоне небольшой рассинхронизации развития зародышей в группе. Использование светофильтров 546 нм и 455 нм, наоборот, тормозило последующее развитие зародышей. Среди эмбрионов, помещенных в эксперимент на стадии поздней бластулы, аномальное развитие наблюдали в группах, разделенных интерференционными светофильтрами с полосами пропускания в более коротковолновой части спектра (372, 379 и 386 нм). После оптического взаимодействия одновозрастных зародышей на стадии поздней бластулы через светофильтр с максимумом пропускания 406 нм наблюдали эффект ускорения эмбрионального развития по сравнению с контрольными группами К2 (в отдельно стоящих кюветах) и К1 (при прямом оптическом совмещении 2-х кювет). Интересно, что узкие спектральные зоны, вычлененные из общего потока сверхслабого биоизлучения, имеют коррегирующее влияние на развитие первоначально одностадийных зародышей, тогда как суммарный поток биоизлучения, включающий те же зоны, таковым не обладает. Это может быть связано с тем, что исключительная стимуляция процессов, чувствительных к определенной длине волны, приводит к нарушению баланса функциональных систем в развивающемся организме. Эксперименты с одностадийными (одновозрастными) эмбрионами показали, что вычленение узких спектральных зон от ближнего ультрафиолета до инфракрасной части оптического диапазона в биоизлучении ранних зародышей в период дробления имеет негативные последствия. На более поздних стадиях проявляется специфичность биологической реакции на определенные спектральные полосы – область 406, а затем и 401 нм (фиолетовая зона) обладает положительным коррегирующим действием, а ближний ультрафиолет (372, 379 и 386 нм) усиливает появление нарушений морфогенеза в группе. И эти стадии соответствуют подготовке к гаструляции – первой этапной дифференциации в самоорганизующейся системе, которой и является развивающийся организм, хотя специфическая активность генома появляется на более ранних стадиях, не чувствительных к спектральной специфичности биоизлучений.
2.3. Изменение параметров сверхслабого излучения эмбрионов низших позвоночных при зеркальном отражении.
Исследования, проведенные с использованием зеркал [45], изготовленных из различных материалов (стеклянных с амальгамовым покрытием, алюминиевых, германиевых и кремниевых), по влиянию отраженного собственного излучения на развивающихся зародышей вьюна показали, что параметры развития эмбрионов после прямого оптического контакта одновозрастных групп (К2) не отличались от контрольных (не имеющих оптического контакта между группами и с зеркалами - К1), но при применении зеркал наблюдали отчетливый аутооптический эффект (изменение скорости дальнейшего эмбриогенеза и появление определенных аномалий развития), причем выраженность аутооптической реакции эмбрионов зависела от материала, из которого изготовлены зеркала, и от схемы пространственного их расположения относительно исследуемой группы эмбрионов. Наибольший биологический эффект наблюдали в случае размещения зеркал одновременно над и под кюветой с эмбрионами. Результаты экспериментов при размещении зеркал только либо под, либо над кюветой с эмбрионами слабо различались, хотя более четкий эффект наблюдали при размещении зеркал под кюветой. Т.е. прохождение излучения по большему пути через несколько сред (вода-воздух-кварц кюветы-зеркало и обратно) при размещении зеркала над кюветой мало меняло результаты эксперимента по сравнению с экспериментами, когда излучение проходит через кварц кюветы, и попадает сразу на зеркало. Проведенные эксперименты могут свидетельствовать о том, что отраженное от зеркал излучение биологического объекта возвращается обратно к нему с некоторым изменением оптических параметров, зависящих в основном от структуры материала и геометрии отражающей поверхности зеркал. Германиевые зеркала «ускоряли» последующее развитие эмбрионов, алюминиевые лишь слабо «ускоряли», тогда как стеклянные – незначительно «замедляли». Наиболее значительный аутооптический эффект, имеющий отрицательные последствия для дальнейшего развития эмбрионов, оказывали кремниевые зеркала. Они не только резко «замедляли», но и обусловливали появление ряда аномалий в дальнейшем развитии до 22 % эмбрионов. В экспериментах использовали два варианта спектрально избирательных кремниевых зеркал. В видимой области спектра вариации спектральных характеристик полупроводникового зеркала достигаются использованием диэлектрических покрытий. При этом за счет высокотемпературного окисления кремния на поверхности пластины создается тонкий слой окисла SiO2 толщиной 0,1-0,3 мкм, прозрачной в области спектра 0,12-7,0 мкм (толщина слоя SiO2 зависит от температуры и времени окисления). В соответствие с разной толщиной диэлектрического покрытия использованные нами кремниевые зеркала имели максимум отражения при l= 520 нм (зеленое зеркало) и l= 650 нм (красно-фиолетовое зеркало). Это позволяло менять спектральный состав возвращенного излучения. Наибольший биологический эффект (замедления скорости развития и появления аномалий) отмечен при использовании зеленых зеркал.
Полученные результаты открывают некоторые подходы к пониманию механизмов аутооптической реакции эмбрионов и к возможностям создания управляемого аутооптического воздействия путем целенаправленной регуляции геометрических параметров зеркального оптического резонатора, применения специальных отражающих покрытий.
2.4.Использование волоконной оптики при исследовании дистантных взаимовлияний развивающихся зародышей низших позвоночных
Установлено, что наибольший биологический эффект оптических контактов биологических объектов наблюдается в кварцевых кюветах, тогда как через обычное лабораторное стекло дистантное взаимовлияние проявляется в очень слабой степени. Дистантное взаимовлияние эмбрионов низших позвоночных активно осуществляется при прохождении границ разных сред: эмбрион-вода-воздух-кварц-вода- эмбрион [46,13]. Вместе с тем, хорошо известно, что эффективность биоизлучения в воздушной среде быстро ослабевает с расстоянием. Так, биологическое действие митогенетического излучения корешков лука эффективно при удалении образцов на расстояние до 1 см, а с увеличением расстояния эффект быстро исчезает [1]. Дистантное взаимодействие разных образцов культуры клеток или бактерий может осуществляться на расстоянии более чуть более 1-2 см [5]. Мы установили, что наибольший эффект взаимовлияния групп разновозрастных эмбрионов, помещенных в отдельные закрытые наполовину заполненные водой стандартные спектрофотометрические кварцевые кюветы, проявляется на расстоянии 1-1,5 см. При увеличении дистанции до 2 см эффективность дистантного взаимодействия, оцениваемая по изменению скорости эмбрионального развития и жизнеспособности (смертности) эмбрионов, снижается на 20 – 25%. При увеличении дистанции между взаимодействующими группами эмбрионов до 3 и 4 см эффективность дистантного взаимодействия снижается соответственно на 60-70 и 80-90%. При расположении групп разновозрастных эмбрионов на расстоянии более 5 см достоверного изменения скорости эмбрионального развития или смертности зародышей по сравнению с соответствующими контрольными группами не наблюдается. Мы исследовали возможность использования волноводов для увеличения дистанции между взаимодействующими группами разновозрастных эмбрионов вьюна [47]. Выявлено, что по металлическим волноводам с отражающими стенками на расстояние более 5-7 см дистантное влияние не распространяется. В отличие от этого, использование различных световодов позволяет передавать эффект дистантного взаимовлияния групп разновозрастных зародышей на более значительные расстояния. Эффективность передачи дистантного взаимодействия биологических объектов зависела прежде всего от типа световода. Выявлено, что 1,5 метровый световод (не передающий при этом четкое изображение объекта) диаметром 7 мм передает эффект дистантного взаимодействия эмбрионов с резким искажением характера биологической реакции. Так, при прямом дистантном взаимодействии групп зародышей 0-й и 6-й стадий развития (на расстоянии 1 см) наблюдается значительное замедление скорости развития и достоверное увеличением смертности эмбрионов младшей возрастной группы по сравнению с контролем, в то время как при разделении кварцевых кювет с зародышами данным световодом наблюдается, наоборот, легкое ускорение эмбрионального развития младшей возрастной группы по сравнению с соответствующим контролем.
Разделение кварцевых кювет с зародышами 0,2 метровым световодом, набранным из тонких стеклянных волокон и имеющим сечение 4 х 4 см (передающий четкое изображение объекта), приводило к передачи дистантного взаимодействия тех же групп эмбрионов без какого-либо искажения характера биологической реакции. При этом как в опыте, так и в контроле в младшей возрастной группе наблюдается значительное замедление скорости развития и достоверное увеличение гибели эмбрионов по сравнению с соответствующими контрольными группами.
Таким образом, использование волоконной оптики позволяет резко увеличивать дистанцию между взаимодействующими биологическими объектами, что позволяет в дальнейшем оптимизировать аппаратуру для применения в практических целях.
ЗАКЛЮЧЕНИЕ.
В настоящее время биология стоит на пороге исследования более глубокого уровня биологических систем – корпускулярно-волновой их организации. Само функционирование биосистемы как результат обмена биосигналами предполагает два взаимно коррелированных уровня этого обмена – вещественный (корпускулярный) и волновой. Вещественный уровень хорошо изучен (матричное копирование ДНК-РНК-белки, механизмы клеточной рецепции, перемещение и самосборка клеточных структур и пр.), а тесно связанный с ним волновой уровень практически не исследован. Все больше появляется свидетельств, что сверхслабые биоизлучения часто оказываются чрезвычано действенным фактором, регулирующим работу биологических систем разного уровня организации. Косвенным доказательством волновой природы описанного эффекта оптической коррекции развития может служить появление эффекта в группах одностадийных зародышей при оптических контактах через устройства, меняющие оптические параметры световых потоков.
Помимо общей коррекции развития результатом дистантного взаимовлияния может быть появление различных аномалий развития, специфических для каждого случая подбора взаимодействующих стадий и оптических условий волновой коммуникации. Особенно ярко это проявляется в нарушениях морфогенезов при формировании головных и туловищных структур зародыша. Так, оптическое совмещение оплодотворенной икры вьюна, приступающей к дроблению, с зародышами, переходящими к гаструляции (т.е. первой дифференцировке на зародышевые листки, дающие в последствии определенные спектры дальнейших дифференцировок органов и тканей) вызывало появление зародышей с раздвоением головного конца (до 8% зародышей), причем уровень раздвоения мог быть в разной степени смещен к каудальной (хвостовой) области зародыша в зависимости от условий эксперимента. Такие аномальные зародыши впоследствии формировали двухголовых предличинок (см. рис.3) с развитыми симметрично или асимметрично (рис.17, Б) головными структурами, с самостоятельными сердцами, работающими в одном или в различающихся ритмах. Однако, если дистантное взаимодействие зародышей тех же стадий осуществлялось при изменении оптических параметров биоизлучения, характер аномалий морфогенеза изменялся.Например, помещение желтого светофильтра в сочетании с поляризатором между кюветами с контактирующими
|
|
|
|
|
А |
Б |
В |
|
Рис. 17. Разные типы нарушений морфогенеза осевого комплекса у вьюна в экспериментах по оптическим контактам. А - предличинка–«шестиголовик» (см. в тексте); Б – две неравномерно развитые головы; В – локальное расхождение осевого скелета на два в задней половине туловища с последующим соединением в хвостовой части. |
||


группами тех же стадий приводило к появлению “двухголовых” зародышей в младшей из взаимодействовавших групп. В старшей группе подобные аномалии не встречались, но появлялись единичные особи с многоголовостью (3–6 голов, разной степени развития) – такой тип аномалий уникален (рис.17,А). Причем в случае, представленном на иллюстрации, головной отдел расчленился на 2 части, каждая из которых образовала по 3 головных структуры малого объема со своими глазами. У этого зародыша в промежутке между двумя комплексами голов образовались 2 сердца с разными ритмами сердцебиений. Изменение соотношения размеров головной и туловищной части развивающихся зародышей мы наблюдали при изменении оптических свойств волновых взаимодействий между эмбрионами тех же стадий (0-й и 8-й) при помещении между кюветами с эмбрионами поляризаторов или интерференционных светофильтров с определенной полосой пропускания. Следует отметить, что оптический контакт зародышей шпорцевой лягушки, проявившей себя наиболее чувствительной к оптическим взаимодейстивиям среди исследованных нами амфибий, так же приводил в определенном проценте случаев к нарушению морфогенеза головных (до появления двухголовости), туловищных и хвостовых структур при включении в опыт зародышей в те же периоды развития, что и вьюны.
Интересно, что некоторые типы морфогенетических аномалий, возникающих в результате дистантного волнового взаимодействия, наблюдались и при экспериментально-эмбриологических исследованиях. Например, микрохирургическое удаление клеточного материала в определенной области зародыша вьюна на 8-й стадии развития приводило к морфогенетическим аномалиям [48], подобным нарушениям, наблюдаемым нами как специфический эффект оптического контакта нормальных зародышей 5-й и 9-й стадий - развивались зародыши без хвостовых и с большим или меньшим недоразвитием туловищных структур (см. рис.5). Дополнительные головные и туловищные структуры можно получить трансплантацией определенного клеточного материала (т.н. первичного организатора), взятого у одного зародыша, в такую область другого зародыша, где в норме они не развиваются, что было установлено классическими экспериментально-эмбриологическими исследованиями. В настоящее время выявлены молекулярно-генетические механизмы, обусловливающие развитие передне-головного отдела зародыша позвоночных – активация работы определенных генов (гомеобоксных группы ANF) в клетках локализованных на определенных участках внешнего зародышевого листка [49, 50]. На формирование пространственного паттерна экспрессии этих генов оказывают влияние клетки другого зародышевого листка - мезодермы [49]. Образование дополнительных осевых структур у зародыша вызывали эктопической экспрессией самих этих генов или введением продуктов их деятельности в необычный для нормального развития участок зародыша [50].
Таким образом, волновые дистантные взаимодействия способны вызывать в развивающемся зародыше процессы эквивалентные либо эктопической активации экспрессии ключевых генов-регуляторов, либо удалению компетентного клеточного материала, отсутствие которого у неповрежденного развивающегося зародыша можно имитировать только блокированием специфического механизма приобретения компетенции. Биоизлучение одних зародышей, специфичность которого подобрана определенным выбором действующей стадии и усилена изменением его оптических параметров (спектральных характеристик, сдвига фаз, поляризации), вызывает морфогенетические аномалии при развитии других зародышей после оптического контакта, вероятно, используя именно эти механизмы. К тому же механизмом, осуществляющим неспецифическую эпигеномную регуляцию, является регуляция уровня свободнорадикальных реакций (одного из параметров окислительно-восстановительного гомеостаза) в развивающемся зародыше, в свою очередь четко реагирующего на биоизлучение. Исследования свободнорадикального механизма реализации воздействия биоизлучения чрезвычайно перспективно, поскольку однонаправленное воздействие может реализовываться либо как стимуляция биохимических процессов в организме, либо имеет цитотоксическое действие при превышении допустимых уровней СР. Было показано, что в развитии организма локальное повышение уровня СР, опосредующее скорость энергетического обмена, соответствует приобретению компетенции эмбриональными зачатками, что является начальным этапом специфических дифференцировок [36]. Полученные нами результаты свидетельствуют, что изменение уровня СР-реакций в зародышах на начальных этапах оптического контакта коррелирует с проявлением дальнейшего биологического эффекта. К тому же, установлено, что наибольшая чувствительность зародыша и его зачатков к повреждающим факторам (в т.ч. и рентгеновскому и лазерному облучению) характеризует именно группы детерминирующихся клеток в развивающемся организме, отличающиеся в норме повышенным уровнем радикалообразования [36]. Вероятно, это в значительной мере объясняет стадиоспецифичность биологического эффекта оптических контактов зародышей.
Т.о. постепенно намечаются молекулярно-биологические (корпускулярные) пути, по которым внешняя волновая информация биологической природы может реализоваться в процессе самоорганизации биосистемы. Это может прояснить роль биоволновой коммуникации внутри самого зародыша при коррекции активации или блокирования определенных подсистем в развитии организма. И если такие биоизлучения способны вызывать морфологические отклонения развития при дистантных взаимовлияниях разновозрастных зародышей, то их участие в регуляции и возможной направленной гармонизации физиологических процессов в самом организме тем более возможно. Сверхслабые информационно-значимые биоизлучения в организме могут иметь регуляторный смысл, особенно, по-видимому, на эмбриональных стадиях развития, играя роль не только сигнального механизма, запускающего различные биологические процессы, но даже определяя во многих случаях их последовательность и, следовательно, направление развития.
ЛИТЕРАТУРА
- Гурвич А.Г., Гурвич Л.Д. Митогенетическое излучение. М.: Медгиз, 1945.- 284c.
2. Magrou J., Magrou M., Reiss P. Actions a distance sur le developpement de l’oeuf d’Oursin. Essai d’interpretation // C.r.s.Acad.Sci.- 1931.-V.193.- P.609-612
- Magrou J. Action a distance et embryogenese // Radiobiologia.- 1932.-N 1. - P.32-38.
- Magrou J., Magrou M., Roubaud M.E.. Actions stimulante ά distance, exercėe par certaines suspensions bactėriennes, ά travers le quartz, sur l`ėclosion du moustique de la fiėvre jaune. C.r.s. Acad. Sci.- 1931. V.192.-P.1134.
- Казначеев В.П., Михайлова Л.П. Сверхслабые излучения в межклеточных взаимодействиях - Новосибирск.: Наука..1981- 144c.
6. Кузин А.М. Роль природного радиоактивного фона и вторичного б иогенного излучения в явлении жизни.- М.: Наука 2002.- 79с.
- Nikolaev Yu..A. Distant interaction in bacterium Pseudomonas fluorescens as a factor of adhesionregulation.// Biophotonics and Coherent Systems. Proc. 2nd A. Gurwitsch Conf. Add. Contributions (Ed.: L. Beloussov, F.-A. Popp, V. Voeikov, R.van Wijk). M.: Moscow University Press. 2000. - P.259-264.
- Николаев Ю.А. Дистантные взаимодействия между клетками бактерий // Микробиология.- 1992/ - Т.61.- №.6.- С.1065-1071.
- Wainwright M., Killham K., Russel C., Grayston J. Partial evidence for the existence of mitogenetic radiation // Microbiology.- 1997.-V.143.- №1.- P.1-3.
- Trushin M.V. Studies on distant regulation of bacterial growth and light emission // Microbiology.- 2003(a).- V.149.- №2.- Р.363-368.
- Trushin M.V. Culture-to-culture physical interactions causes the alteration in red and infrared Escherichia coli light stimulation of growth rate // Journal of Microbiology, Immunology and Infection.- 2003(b).- V.36.- P.149-152.
- Белоусов Л.В., Попп Ф.-А., Казакова Н.И. Сверхслабые излучения куриных яиц и зародышей: неаддитивное взаимодействие двух излучателей и устойчивая неравновесность //Онтогенез. -1997.-Т.28.- N5. - С. 377-382.
- Burlakov A.B.Distant physical interactions between the developing fish embryos// Biophotonics and Coherent Systems. Proc. 2nd A.Gurwitsch Conf. Add. Contributions (Ed.: L. Beloussov, F.-A.Popp,V. Voeikov, R.van Wijk.) M.: Moscow University Press.-2000.-P.289-304
- Moltchanov A.A., Galantsev V.P. On the functional role of spontaneous photon emission in the mammary gland // Biophotonics. Non-equilibrium and coherent systems in biology, biophysics, biotechnology. Proc.Intern. A.G.Gurwitsch conf. BioInform Services M.: Moscow University Press . -1995. - P.341-350.
- Novikov K.N., Voeikov V.L., Popp F.-A.. Analysis of light emission by neutrophils in the process of respiratory burst suggess that physical fields are involved in intercellular communications. // Biophotonics. Non-equilibrium and coherent systems in biology, biophysics, biotechnology. Proc. Intern. A.G.Gurwitsch conf. BioInform Services M.: Moscow University Press .- 1995.- P.291-302
- Xun Shen, Wei Ping Mei, Xing Xu. Information transfer from the neutrophils undergoing respiratory burst to second chemically separated but optically coupled neutrophil population. // Biophotonics. Non-equilibrium and coherent systems in biology, biophysics, biotechnology. Proc. Intern. A.G.Gurwitsch conf. BioInform Services M.: Moscow University Press. - 1995. - P.303-316
- Николаев Ю.А. Регуляция адгезии у бактерий Pseudomonas fluorescens под влиянием дистантных межклеточных взаимодействий // Микробиология.- 2000. - Т.69.-№.3.- С.356-361.
- Beloussov L.V., Burlakov A.B., Konradov A.A. Biophoton Emission from Eggs and Embryos of a Fish, Misgurnus fossilis L.: Developmental Dynamics, Frequency Patterns and Non-Additive Interactions //Biophotonics and Coherent Systems. Proceedings of the 2nd Alexandr Gurwitsch Conference and Additional Contributions (Editors: Lev Beloussov, Fritz-Albert Popp, Vladimir Voeikov, and Roeland van Wijk). Moscow University Press, Moscow.- 2000. - P.305-320.
- Белоусов Л.В., Бурлаков А.Б., Лучинская Н.Н. Статистические и частотно-амплитудные характеристики сверхслабых излучений яйцеклеток и зародышей вьюна в норме и при их оптических взаимодействиях. I. Характеристики сверхслабых излучений в нормальном развитии и оптическая роль яйцевых оболочек. // Онтогенез. 2002.- Т. 33. - № 3.- С. 213-221.
20. Белоусов Л.В., Бурлаков, А.Б., Лучинская, Н.Н. Статистические и частотно-амплитудные характеристики сверхслабых излучений яйцеклеток и зародышей вьюна в норме и при их оптических взаимодействиях. II. Изменение характеристик сверхслабых излучений при оптическом взаимодействии разновозрастных групп зародышей. // Онтогенез. - 2003.- Т. 34.- № 6. -С. 453-463.
- Бурлаков А.Б., Бурлакова О.В., Голиченков В.А. Дистантные взаимодействия разновозрастных эмбрионов вьюна // ДАН. -1999.- Т.368.- N4. -С.562-563.
- Бурлаков А.Б., Бурлакова О.В., Королев Ю.Н., Голиченков В.А. Самоорганизация развивающихся организмов и поляризация волновой биокоммуникации // Синергетика.- Т.3.- М.: МГУ. 2000. -С.296-307.
- Бурлаков А.Б. Бурлакова О.В., Голиченков В.А. Дистантные волновые взаимодействия в раннем эмбриогенезе вьюна Misgurnus fossilis L // Онтогенез.- 2000.-Т.31..- №5..- С.343 – 349.
- Костомарова А.А. Вьюн Misgurnus fossilis L. //Объекты биологии развития. М.:Наука. 1975. - С.308 – 323.
- Дабагян Н.В., Слепцова Л.А. Травяная лягушка Rana temporaria L.// В кн.: Объекты биологии развития. М.: Наука. - 1975.- C. 442-463.
- Cambar R., Marrot B., Table chronologique du developpement de la grenouille agile (Rana dalmatina Bon.). Bull. Boil. France-Belgique. – 1954. -V 88. – P. 168-177.
27. Nieuwkoop P.D., Faber J. 1956.Normal table of Xenopus laevis (Daudin). - Amsterdam, North-Holland Publ. Co..- 295 p.
28. Cambar R., Gipouloux. 1956. Table chronologique du developpement embryonnare et larvaire du crapaud commun Bufo bufo L. Bull. biologique de la France et de la Belgique, XC F2.
- Burlakov A.B., Golichenkov V.A. The long range interactions of developing biosystems // Modern Natural Sciences: Proceedingss of 3rd International Conference INTERNAS’2007, Kaluga, Russia. May 22-25, 2007. –Kaluga: KGPU. – 2007. - .P. 115-118.
- Детлаф Т.А. Температурно-временные закономерности развития пойкилотермных животных. М.: «Наука». 2001. - 211с.
- Доронина Н.Ю., Бурлакова О.В., Бурлаков А.Б., Голиченков В.А Темпы развития зародышей вьюна Misgurnus fossilis L. и проявление дистантных взаимовлияний в раннем онтогенезе // ДАН. -. 2002. - Т.384. -. N1. -.С. 136-138.
- Бурлакова О.В., Доронина Н.Ю., Бурлаков А.Б., Голиченков В.А. К механизмам восприятия информации при волновой биокоммуникации в раннем развитии низших позвоночных // Неионизирующие электромагнитные излучения в биологии и медицине (БИО-ЭМИ-2002). Труды международной конференции. Калуга. 2002.- С. 74-78.
- Бурлаков А.Б., Аносов В.Н., Наумов Н.А., Перевозчиков Н.Ф., Пушкарь В.Я., Голиченков В.А. Использование физических носителей для анализа волновой информации биологических систем // Электромагнитные излучения в биологии (БИО-ЭМИ-2000). Труды международной конференции. (Калуга, Россия, 3-5 октября 2000 года). 2000.- С.46-49.
- 34 Бурлаков А.Б., Аверьянова О.В., Пушкарь В.Я., Голиченков В.А. Информационная роль биофотонной эмиссии при эмбриональном развитии. // Актуальные проблемы современного естествознания. Интернас Калуга:. КГПУ. - 2000. -C.70-71
35. Бурлаков А.Б., Бурлакова О.В., Аносов В.Н., Перевозчиков Н.Ф., Пушкарь В.Я., Голиченков В.А. Модулирование параметров биофотонной эмиссии для анализа волновой информации в саморазвивающихся биологических системах // Синергетика.- М.: МГУ. 2001. - Т.4. - С. 288-299.
36. Мелехова О.П. Свободнорадикальные процессы в пространственно-временной регуляции развития низших позвоночных. Докт.дисс. М.,МГУ. - 2005.
37. Voeikov V.L., Processes involving reactive oxygen species are the major source of structured energy for organizmal biophotonic field pumping. In: Biophotonics and coherent systems. Moscow university press. – 2000. – Р..203-228.
- Козлов Ю.П. Привитая сополимеризация как метод исследования свободных радикалов в биологических системах. М.: изд-во МГУ.- 1970.
- Мелехова О.П. Свободно-радикальные процессы в эмбриогенезе Anura. Онтогенез. - 1976. - Т. 7. - №2. - С. 131-135.
- Бурлакова О.В., Бурлаков А.Б., Малахов Ю.И., Королев Ю.Н., Голиченков В.А. Поляризационные и фазовые эффекты при дистантном взаимодействии биологических объектов // Оптические методы исследования потоков: Труды VI Междунар. научно-технической конф. – М: Изд-во МЭИ. – 2001. -.С.374-377.
- Бурлаков А.Б.,Голиченков В.А, Бурлакова О.В., Королев Ю.Н. Поляризационые эффекты при дистантном взаимодействии биологических объектов Вестник МГУ. Серия 16. Биология. – 2002. - N2 - С.3-8.
- 42 Бурлакова О.В., Бурлаков А.Б., Королев Ю.Н., Голиченков В.А. Волновая биокоммуникация и самоорганизация развивающихся биосистем // Синергетика. -2000. .Т.3.- МГУ. М.- С 307-313.
- Бурлаков А.Б., Медведева А.А., Бурлакова О.В., Малахов Ю.И., Голиченков В.А. Особенности биологического действия спектральных составляющих сверхслабых излучений вьюна в раннем онтогенезе // Сборник избранных трудов IV международного Конгресса "Слабые и сверхслабые поля и излучения в биологии и медицине”. (Санкт-Петербург 03-07.07.2006).- 2006.- С.34-40.
- Бурлаков А.Б., Малахов Ю.И., Медведева А.А., Бурлакова О.В., Голиченков В.А. Анализ спектрального состава сверхслабых излучений ранних зародышей вьюна // Оптические методы исследования потоков: Труды IX Международной конф. – М.: Издательский дом МЭИ.- 2007. - С. 328-331.
- Бурлаков А.Б., Шарихин В.Ф., Бурлакова О.В., Голиченков В.А. Изменение параметров сверхслабого биоизлучения эмбрионов низших позвоночных при зеркальном отражении// Электромагнитные излучения в биологии. Труды III международной конференции. Калуга, Россия, 5-7октября 2005. Калуга.: КГПУ. - 2005. - С. 47-51.
- Бурлаков А.Б. Внутривидовая волновая коррекция раннего эмбрионального развития //Пространственно-временная организация онтогенеза (под ред. Ю.А.Романова, В.А.Голиченкова) - М.: МГУ..- 1998..- С. 183— 188.
- Бурлаков А.Б., Малахов Ю.И., Перевозчиков Н.Ф., Шарихин В.Ф., Голиченков В.А. Использование волоконной оптики при исследовании дистантных взаимодействий биологических объектов // Оптические методы исследования потоков: Труды YI Международной научно-технической конф. (под ред. Ю.Н. Дубнищева, Б.С. Ринкевичюса). – М.: Изд-во МЭИ. – 2001. - С. 380-384.
- Слепцова Л.А., Сильченко Д.А., Неклюдова И.В., Корвин-Павловская Е.Г., Голиченков В.А. Исследование регуляционных способостей бластулы вьюна и потенций клеток дорсальной области бластодермы // Онтогенез. -1999.- Т. 30.- N 2.- С.97-102.
- Kazanskaya O.V., Severtzova E.A., Barth K.A., Ermakova G.V., Lukyanov S.A., Benyumov A.O., Pannese M., Boncinelli E., Wilson S.W., Zaraisky A.G. ANF: a novel class of homeobox genes expressed at the anterior end of the main embryonic axis of vertebrates. // Gene. - 1997.-V.200.- P.25-34.
- Зарайский А.Г.. Гомеобоксные гены класса ANF –регуляторы раннего развития передне-головного отдела у позвоночных. // Автореф. докт.дисс., М. 2000