A FRACTAL SPACE-TIME
STRUCTURE OF AN OPEN LIVING SYSTEM
Victor Gusev
Sobolev Institute
of Mathematics, Siberian Branch, Russian Academy of Sciences, Novosibirsk, Novosibirsk State
University, Novosibirsk, Russia
The experimental data described in papers [1–24] are
theoretically analyzed to construct an adequate model for dynamics of an open nonequilibrium living system. In addition, we have
attempted to provide a generalized description of the properties of living and
nonliving matter. Relevant published data were used to demonstrate a fractal
structure of the space in the vicinity of centrally gravitating bodies with
satellites revolving around them along closed trajectories and serving as a
kind of testers of the neighboring space. A local violation of its
discontinuity is likely to be a necessary (yet not sufficient!) dynamic
characteristic of the spatiotemporal continuum for self-organization of
molecules into a living, i.e., self-replicating, system.
INTRODUCTION
Over 50 years ago, two
Thirty years ago, our research team
by a lucky confluence of circumstances discovered an analogous phenomenon of
reproduction of an organotrophic microorganism, E. coli,
under even more stringent conditions, in tetradistilled
water. These experiments are described in [3–7]. There we also formulated a
biophysical model for this phenomenon. Additional experimental data on the
evolutionary drift of the number of viable cells
(hereinafter, NVC), cooperative effects, quasiperiodic
variations of the mean NVC value in a sample, as well as relative variance and
biorhythms in the microbial communities under extreme conditions of absolute
substrate starvation, i.e., in a superpure distilled
water, are briefed in [8]. Below, we describe the results of further studies
into the properties of microbial populations in a substrate-free medium.
Nonlinear Phenomena in Microbial
Population Dynamics
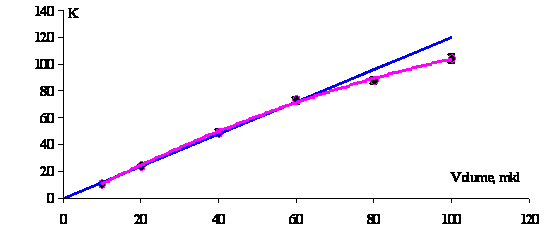
Figure 1 shows the dependences
of NVC on the sampling volume taken from the initial population. The initially
linear dependence
for small sampled volumes deviates from a linear pattern in a statistically significant
manner with increase in this volume. The value and pattern of such deviations
are not strictly fixed for an individual sample in population yet change with
time (see Fig. 6).
|
|
|
Fig. 1. Nonlinear dependence
of the number of viable cells in sampled volume. The ordinate shows the NVC in sample. |
Consider one more
nonlinear effect associated with a change in concentration of microbial
suspension with its dilution with water (Fig. 2). It could be a priori
expected that an m-fold dilution of the suspension containing K0
cells per unit volume will change the cell as
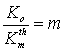 ,
(1)
,
(1)
where denominator is the theoretically expected value of
microbial concentration. However, the experimental data transform this
dependence to
![]() ,
(2)
,
(2)
where denominator is the experimentally
observed microbial concentration.
|
|
|
Fig. 2. Nonlinear dependence of concentration on
dilution factor. Nonlinearity coefficient α = 0.783 ± 0.023;
correlation coefficient of nonlinear approximation r = 0.999; and
solid line, linear approximation according to initial data. |

The nonlinearity
coefficient α is not a strictly fixed constant. Its value is mainly
determined by the time of relaxation to stationary state after mechanical
disturbance of population (see Figs. 4 and 6).
Figure 3 shows the
experimental dependence
of a change in NVC after dilution of the initial concentrated suspension (K ≈ 108
cm–3) to the concentration K ≈ 103
cm–3. Within the error, the parameters of relaxation curve are
independent of the degree of population dilution.
Note that immediately
after dilution of cell suspension, NVC drops strictly twofold independently of
the dilution factor and then again relaxes to the initial concentration.
Evidently, there is no question of any irreversible mechanical destruction of
half cell population. Presumably, after a mechanical disturbance of population,
a rapid destruction of autonomously stable spatial population structure takes
place with its subsequent slow relaxation to the state that provides all cells
with the amount of free environmental energy sufficient for maintaining the
viability of the entire population.
The experimental data suggest
that
·
Nonlinear phenomena are observed when sampling the population
with a constant cell concentration, namely, the NVC in samples increases not
directly proportionally to their volume;
·
The change in NVC is also not directly proportional to the
dilution factor after changing cell concentration (diluting cell suspension via
addition of water); and
·
The spatial structure of population is rather sensitive to
external mechanical disturbance but is able to relax to the initial state.
|
|
|
|
|
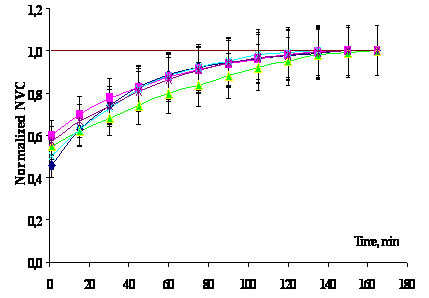
Fig. 3.
Relaxation of the number of viable cells (NVC) in water at a room temperature
after dilution of the initial suspension. The initial population with a
concentration of viable cells of ~108 cm–3
was kept at +4ºC for 2 days to immediately (for 5–7 min) prepare five
tenfold dilutions (initial suspension with 108 cm–3;
next series 107 cm–3, 106 cm–3,
105 cm–3, 104 cm–3 and 103
cm–3) with distilled water at a room temperature and sample them
periodically for determining NVC. The NVC value (the ordinate) was normalized
by dividing the NVC at a given moment by stationary NVC value (upper
curves). Averaging was performed over all dilutions (see upper curves).
The approximation curve K/K0 = 1 – 0.5exp(–t/42.7)
with the correlation coefficient r = 0.8; relaxation time |
To determine the physical meaning of
nonlinearity coefficients, we use fractal geometry methods. As is known, the
objects of fractal geometry are the structures with a characteristic hierarchy
and a scale invariance.
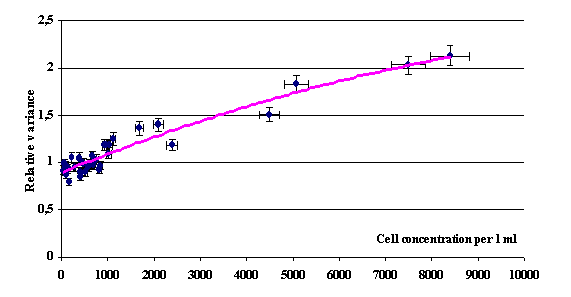
See Fig. 4, which shows the dependence of the relative variance ‹s2/![]() ›T
averaged over time at various cell concentrations (the mean NVC value in sample
›T
averaged over time at various cell concentrations (the mean NVC value in sample
![]() is proportional
to the mean cell concentration
is proportional
to the mean cell concentration ![]() , because the samples
of strictly fixed volume were taken; s , the variance in a series of samples).
The time intervals of averaging were changed in a random manner from point to
point in the range of 1–4 h. This processing method was selected to level the
accuracy error inevitably introduced when counting NVC.
, because the samples
of strictly fixed volume were taken; s , the variance in a series of samples).
The time intervals of averaging were changed in a random manner from point to
point in the range of 1–4 h. This processing method was selected to level the
accuracy error inevitably introduced when counting NVC.
These data suggest that a cell concentration
of 8 × 102 cm–3 is the critical point
below which the variance s2/![]() < 1; with increase in cell concentration, the
relative variance exceeds unity and even permanently grows. This effect can be
interpreted as a transition from quasi-regulated distribution of viable cells
at concentrations below 8 × 102 cm–3 in
the volume to heterogeneous distribution, when a hierarchy of clusters with
different concentrations of viable cells is formed. In addition, a complex
evolution of these clusters is observed. This conclusion was made based on the
variations of relative variance around the mean value with time; this mean
value is determined by the concentration of viable cells (see Figs. 5 and
< 1; with increase in cell concentration, the
relative variance exceeds unity and even permanently grows. This effect can be
interpreted as a transition from quasi-regulated distribution of viable cells
at concentrations below 8 × 102 cm–3 in
the volume to heterogeneous distribution, when a hierarchy of clusters with
different concentrations of viable cells is formed. In addition, a complex
evolution of these clusters is observed. This conclusion was made based on the
variations of relative variance around the mean value with time; this mean
value is determined by the concentration of viable cells (see Figs. 5 and
|
|||
|
Fig. 4. Dependence of relative variance s2/ |
Without getting into the physical
mechanism of this phenomenon, consider its formal aspect. In the case of a
hierarchical self-similar distribution of elementary units forming a fractal
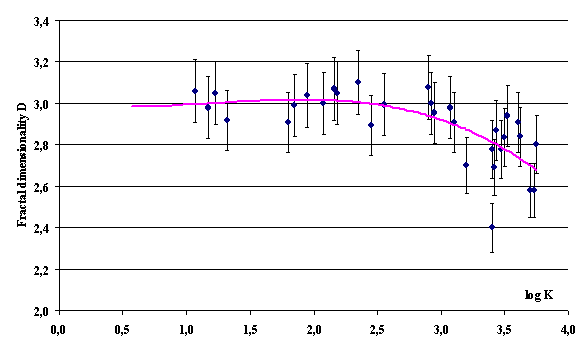
object in three-dimensional Euclidean space, their density in a volume is not a
constant but rather depends on the sample volume as follows:
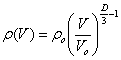 ,
(4)
,
(4)
where r(V) is the density of elementary units in the sampled volume V
(in this case, NVC); ro, average
density over the entire volume occupied by fractal object; Vo,
the volume of one elementary unit (in this case, the volume per one cell, a
uniform distribution of the overall population provided); and DR,
fractal dimensionality of the object. It follows from Eq. (4) that at m-fold
dilution of the population displaying fractal properties, the corresponding
densities r(V) and rm(V) are related with one another as
![]() .
(5)
.
(5)
As it follows
from Eqs. (2) and (5), the fractal dimensionality and
nonlinearity coefficient are related as
D = 3α
(6)
Figure 5 shows the dependence of fractal dimensionality on the concentration of microorganisms
calculated using this equation.
At a cell concentration exceeding 3 × 103 cm–3,
the curve in figure commences significantly deviating from asymptotic
value of 3, characteristic of small concentrations.
Relaxation
Dynamics of Fractal Dimensionality
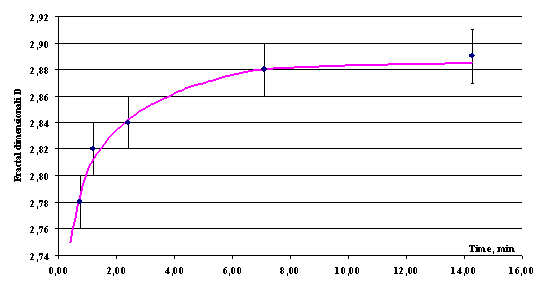
Figure 6 shows the relaxation curve
for fractal dimensionality of microbial population after mechanical
disturbance.
|
|
|
Fig. 5. Dependence of fractal dimensionality on the
concentration of viable cells. The abscissa shows the decimal logarithm of
cell concentration per 1 cm3. |
|
|
|
Fig. 6. Dependence of the fractal dimensionality of population on the relaxation time after mechanical disturbance. Concentration of suspension is (3.5 ± 0.07)× 103 cm–3. |
The essence of experiment was as follows: suspension
of microbial cells was intensively stirred and then, as soon as possible (as a
rule, over 15–20 s), 5 ml of this mechanically disturbed suspension was pumped
using a peristaltic pump into a separate glass container (tube). Then samples
were taken from this tube and multiply diluted with water (as a rule with a
step of two). The diluted samples were plated onto petri
dishes containing agar nutrient medium to determine the concentration of viable
cells. The dishes were incubated overnight at 37ºC to count the colonies
on the next day and determine the fractal dimensionality using Eq. (6).
As is evident from Fig. 6, the mechanically
disturbed population completely relaxed to the stationary value of fractal
structure over more than 15 min. It was necessary to take this fact into
account when determining the dimensionality D for the population exposed
to mechanical disturbance, for example, after dilution.
To construct a self-consistent spatiotemporal
model for living systems, we then need the experimental data on the time drift
and dynamics of microbial populations. As has been demonstrated [3–8], it is
almost impossible to observe a living system in a stationary state. For the
observation times from seconds to several days when the population of organotrophic microorganisms in a substrate-free medium was
monitored, the NVC permanently varied by tens of percent from the mean value
(for small times) to several orders of magnitude and back to the mean value at
long times (see Figs. 1–3 in [8]). In other words, it is also possible to
introduce the concept of a time fractal for populations of the living systems
that form a spatial fractal structure [10].
A Fractal Structure of Space and Time in
Living Systems
In both the everyday live and
scientific research, it is a priori considered that the space where we
live has integer dimensionalities: line is one-dimensional, plane is
two-dimensional, and space has only three dimensions. It seems that it is
impossible to imagine the world and the objects in it with fractional
dimensionalities. However, this is a mere consequence of our conservatism and
habits. A coastline, clouds, porous structures, electrical discharges in
atmosphere, and many other natural phenomena are much easier describable with
the help of the language of mathematical equations by introducing the concept
of fractality, i.e., a fractional dimensionality. The
branched network of blood vessels of warm-blooded animals as well as their
neuronal network, branching structures of trees, and so on can be described in
an analogous manner. It appeared a surprising fact for us that the distribution
of viable aquatic microbial cells in a volume can also be described using the
language of fractals.
Figure 2 shows the
approximation of experimental data with the nonlinearity coefficient α = 0.783 ± 0.023.
These data make it possible to determine the fractal dimensionality of the
set of viable cells distributed in population volume as D = 2.35 ± 0.07.
The fractal dimensionality of time is determined
when analyzing long-term time variations in NVC in microbial population via the
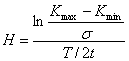 ,
(7)
,
(7)
where Kmax
and Kmin are the maximal and
minimal NVCs in the array of samples taken during the
time T with an interval t and s,
the variance for this array. The number of samples in the array, N = T/t,
in our experiments was no less than 600, because it was demonstrated beforehand
that the Hurst constant in such array reached a stationary value, which could
be later used for calculating the fractal dimensionality of time using the
equation from [10]:
DT
= 2 – H,
(8)
where DT is
the fractal dimensionality of time for the data array.
Figure 7 demonstrates the dependence of the
|
|
|
Fig. 7. Dependence of the |
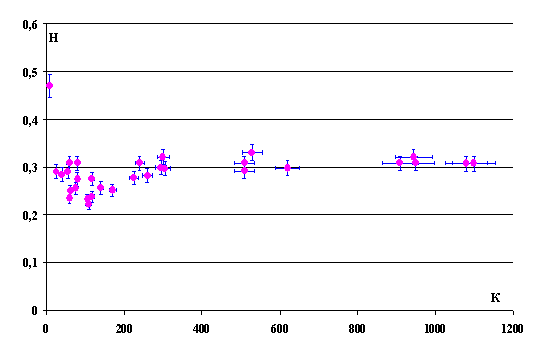
It is easy to calculate the
fractal dimensionality of population, DT
= 2 – Hst(K) = 2 – 0.309 = 1.691 ± 0.004.
A formal description of objects and processes,
accepted in physics, is performed by specifying the corresponding functions
determined in the space of three variables, the coordinates, and the variable
independent of them, time. In relativistic physics, these variables are
regarded as 3 + 1, i.e., four-dimensional, spatiotemporal continuum. In our
case, this sum is represented as D + DT = (2.35 ± 0.07)
+ (1.691 ± 0.004) = 4.04 ± 0.07.
The correctness for such summing of the dimensionalities of fractal sets is
based in the Mandelbrot empirical rule [10]. Of interest is the fact that two
fractional values, spatial and temporal fractal dimensionalities
determined independently and with a high accuracy, give in sum with the same
accuracy the integer dimensionality value equal to the dimensionality of the
spatiotemporal continuum of the Universe.
Similar to all the previous arithmetic and algebraic
cases with living systems [12, 13], this relation is not a proof but only a
hint to researchers at the way where to search for the association between
living and nonliving in the Universe and an indication of their unity.
Fractal
Properties of Dynamic Systems on Cosmic Scales
Modern physics, which is the
foundation for all natural sciences, is unable to explain both the phenomenon
of life itself and the molecular mechanism of the origin of life. i.e., the
conditions required for self-organization of atoms into a self-replicating
system, the elementary unit of which is a microbial cell. In other words, based
on the physical postulates—the conservation laws, which are the corollaries of
the abstract concepts of uniformity and isotropy of space and uniformity of time—does not provide for
construction of a consistent model of the Universe, during the evolution of
which self-replication molecular systems could emerge. The
overall experimental and theoretical pool of knowledge tested so far since the
1920s has given neither positive nor negative answer to the question on the
natural origin of the terrestrial life formulated by Oparin
[20a].
It is possible that such current of
thought of the fathers of modern science was initially determined by the laws
underlying the evolution of the Universe, which thereby tabooed unraveling of
the secret and meaning of all living on the Earth. In an allegoric form, we can
follow the idea of Strugatskie [21] that the
Macrocosm preserves its homeostasis to a certain stage of the evolution of
human civilization. Indeed, everything that has been ever discovered in Nature
by the Civilization “immature enough” to understand the meaning of a discovery
has been first used to destroy this Nature and only later, for certain peaceful
application aimed at a more comfort survival, as being exemplified by
explosives, poisonous gases, and nuclear energy. Having acquired the ability to
create artificial living systems, such “immature” Civilization will inevitable
destroy itself, since an absolute control over self-replicating systems, for
example, pathogenic microorganisms, is currently impossible. There always will
be some individuals with a pathological desire for the control over the globe,
which, as we know from history, has always ended in a catastrophe for both
dictators and the human communities they govern.
The only thing left to scientists
is to debate on whether the origin of the life on Earth was natural or imported
as “seeds” somewhere from the Universe. We can state that a periodic domination
of either of the competing hypotheses—panspermia and
natural chemical evolution—is the result of a technological breakthrough in
either biochemistry or astrophysics. However, neither hypothesis has any
constructive origin, because the carriers of these ideas are the scientists
whose worldview has been formed within the frame of the current science, unable
to explain the phenomenon of life.
Philip Morrison, an American physicist, noted
that the discovery of life on one other planet can “transform the origin
of life from a miracle to a statistic”, that is, to a certain
experimentally reproducible thing, which forms the background for the
methodology of scientific cognition of the world. However, neither physics nor
biology can answer the question on what is the essence of the experiment that
could simulate the conditions of the origin of living systems2.
Appealing to the opinion of the founders of
quantum mechanics, we should admit that a constructive solution for this
problem (i.e., the problem of the origin of life) requires formulation of a
“crazy” hypothesis. This work is an attempt in this direction.
Based on an anthropic
principle [14], we can postulate that the living self-replicating objects must
inevitably emerge at a certain stage in the evolution of the observable
Universe. In other words, the event or process of emergence of simplest life
forms, the evolution of which had led to the origin of the observer himself,
man, is potentially inherent for all the known and, possibly, yet unknown
physical laws. Otherwise, i.e., in the absence of the latter, the Universe is
unobservable and is an abstract notion. The above reasoning suggests that not
only the laws that determine molecular processes, but also the laws of the
Macrocosm are responsible for the origin and evolution of living systems.
Gorelik
in his monograph [16] cited and discussed the position of Ehrenfest
[17], who asked “In what way does it become manifest in the fundamental laws of
physics that space has three dimensions?” or, in other words “What specific
features distinguish the geometry and physics in R3 from the geometries and physics in Rn?”
Thus formulated, these questions are possibly senseless. Undoubtedly, they are
subject to criticism, because this entails the questions on whether the space
“exists” at all, whether it is three-dimensional, and the very questions “why?”
and what “physics” means in the case of R3 or R7?
Gorelik did not try to find a
better wording for these questions. Presumably, others will succeed in
specifying more singular features of R3, and it will then become
clear what correct questions match the answers provided by our considerations
[16].
Ehrenfest
pioneered in formulating the problem of
three-dimensionality of the physical space of the observable Universe at both
micro and macro levels as long ago as 1917 [17]. In his works, he theoretically
justified the fact that the space dimensionality provided, the existence of
stable closed trajectories of interacting objects and their movement to
infinity are possible only when D = 3. This conclusion is true for the objects
interaction via both the electric and gravitational fields, because in either
case the inverse square law is met.
Thus, the following
statement is made: living objects could have appeared only in such Universe
as it shows up to the observer. This statement is actually one of the
simple definitions of the anthropic principle,
introduced into scientific use by Dicke [14].
Further analysis of the
properties of living matter will be based on the following postulates:
· Space
does not exist by itself in the absence of elementary structural units of
nature and
· The
geometry of space, its dimensionality, as well as topological and metrical
characteristics are determined by the pattern of mutual arrangements of
structural units and the laws of their physical interactions.
These definitions
contain nothing principally new and, therefore, contradictory to modern
physics. On the other hand, the definition of space as a derivative of physical
objects and the laws of their interaction allows its dimensionality to be also
considered as a derivative rather than and axiomatic notion. In this case, the
problem of space dimensionality and its integer value D = 3 transits
from axioms to an interpretable category [15, 16]. Depending on the structure
of objects, their scales, and patterns of interaction and mutual arrangement,
the space dimensionality can be strictly integer or take fractional values.
This statement can be illustrated by actual calculations based on astronomic
observations of movements of planets in the Solar system and their satellites.
Commence constructing
the hypothesis with Kepler’s third law:
![]() ,
(9)
,
(9)
where M is
the mass of central gravitating body; T, the period of satellite
revolution around it; R, radius of the major semiaxis
of the orbit, and G, gravitational constant. It is convenient to
represent Eq. (9) in a dimensionless form:
![]() ,
(10)
,
(10)
where Ro
is the radius of the central gravitating body and To, the
period of satellite revolution at this distance. Such nondimensionalization
provides for analyzing both planets and their satellites in a unified system of
coordinates.
Analysis of the astronomic data
for planets of the Solar system and their satellites [22] demonstrates that at r = R/RE < 0.1
3 (in astronomic units, where RE is the distance
between the Earth and Sun), Eq. (10) starts to considerably deviate from unity;
moreover, the variations around unity increase with a decrease in the distance
of satellites from planets. Formally, it should be interpreted so that the laws
of celestial mechanics formulated for a three-dimensional space fail to
precisely describe the movement of bodies at the scales r = R/RE < 0.1
(in astronomical units). For further analysis of this phenomenon, we use the
generalized form of dimensionless Kepler’s Eq. (10)
proposed by Ehrenfest [15] for the space of an
arbitrary dimensionally D:
![]() , from
which it follows that:
, from
which it follows that:
![]() .
(11)
.
(11)
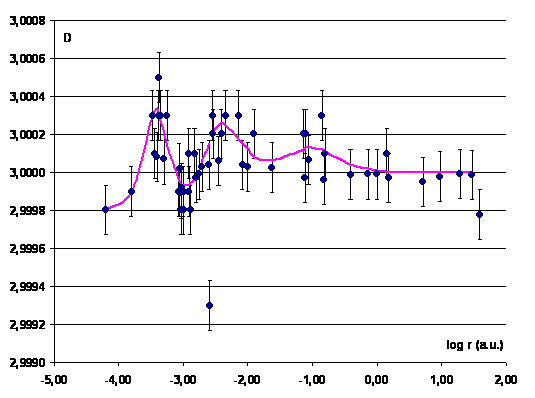
Figure 8 shows the calculation results using
this equation defining the parameter D for the objects
of the Solar system known by the year of 1990 [22, 23]. As is evident from Fig.
9, the amplitude of variation of this parameter increases in a statistically
significant manner with a decrease in the distance. The point D =
2.9993 corresponds to the Moon.
The only object falling out of the entire data
array for satellites is the Moon (log r = –2.59). As for the
planetary scale from the Mercury to
The author does not assume the responsibility
for interpretation of the physical reasons underlying this phenomenon. However,
since the goal of this work is to formulate a hypothesis verifiable for the
origin of life to the extent possible that would fundamentally differ from all
the previously proposed hypotheses, we well use the results obtained by
analyzing variations in the parameter D to formulate
the following hypothesis:
In the vicinity of gravitating
masses in the space on the average uniform and isotropic, spherical layers of a
fractal space structure are formed, the dimensionality of which is subject to
small variations in the neighborhood of the integer value D = 3.
Consider the possible consequences of such
variation in space dimensionality in the context of the stated problem.
Assuming that the observed variation is not a
consequence of an accuracy error (which is actually impossible, because
otherwise it would be similarly impossible to observe the variations in DR
but only a monotonic drift), we can postulate that the physical vacuum
additionally influences the pattern of satellite motion. Vacuum is among the
most mysterious material substances, whose properties are being discussed among
theoretical physicists for already several decades. We will take advantage of
the current concepts on physical vacuum by various authors [18–20] to try to
extract the rational kernels for construction our own view on solution of the
problem of the origin of life.
During the last decade, the terms dark
energy and antigravitation [20] have been
formulated and almost established in the scientific worldview. Although the
concept of antigravitation was introduced by Einstein
as long ago as 1917, it was really comprehended only after analyzing the
cosmological observations during several recent years. In fact, antigravitation significantly manifests itself in repulsion
of galaxies on a scale of five to eight billion light years,
Therefore, it is quite evident that it is fundamentally impossible to observe antigravitation by the same methods on planetary scales to
say nothing about the scale of Earth. However, the goals of this work do not
require this, since the mere fact of antigravitation
and its recognition in the scientific community is sufficient.
CONCLUSIONS
From the standpoint of the author, in addition
to the standard physiologically necessary conditions for sustainable existence
of life, namely, “normal” pressure, temperature, and humidity of environment
and the absence of pathogenic physical and chemical factors, polarization of
the physical vacuum is a basically important factor for self-organization of
molecules into a self-replicating system [18, 19]. The consequence of this is a
fractal, i.e., noninteger, space–time dimensionality,
which entails nonlinearity of all molecular processes eventually giving rise to
a spatiotemporal ordering of the molecular processes that had led to formation
of self-replicating informational structures. Note that the noninteger,
i.e., fractional, dimensionalities of both space and time separately also gave
in sum an integer, an invariant of four!
|
|
|
Fig. 8. Dependence
of the parameter D on the distance of satellite from the
central body. The abscissa shows the decimal logarithm of the distance in
astronomic units. |
Acknowledgments
The author is grateful to Prof. A. D. Gruzdev, Prof. S.A. Shnoll, Prof.
R.G. Khlebopros for fruitful discussions.
REFERENCES
1. Elin, V.L., On the biology of coliform
bacillus, Mikrobiologiya, 1957, vol.
26, issue 1, pp. 17–21 (in Russian).
2. Kalinenko, V.O.,
Reproduction of heterotrophic bacteria in distilled water, Mikrobiologiya,
1957, vol. 26, issue 2, pp. 148–153 (in Russian).
3. Gusev, V.A., On the source of energy for preservation of viability and
amplification of heterotrophic microorganisms under conditions of substrate
starvation. I. Formulation of hypothesis, Biofizika,
2001, vol. 46, issue 5, pp. 862–868 (in
Russian).
4. Gusev, V.A. and
5. Gusev,
V.A., On the source of energy for preservation of
viability and amplification of heterotrophic microorganisms under conditions of
substrate starvation. III. Necessary and sufficient conditions for
microorganisms to colonize water envelopes of planets, Biofizika,
2001, vol. 46, issue 5, pp. 875–878 (in
Russian).
6. Gusev,
V.A., Evolution of microbial cell population under complete substrate
starvation, in: Evolutionary Simulation and Kinetics, Yu.I. Shokin, Ed.,
7. Gusev, V.A., Evdokimov, E.V., and Bobrovskaya,
N.I., Deviation from Poisson distribution in the series of identical E. coli
cell culture, Biofizika, 1992, vol. 37,
issue 4, pp. 733–737 (in Russian).
8. Gusev,
V.A., Evolutionary drift, cooperative effects, quasiperiodic
variations, and biorhythms of microbial communities under extreme conditions of
absolute substrate starvation. Zh. Problem Evol. Otkr. Sistem, 2008, vol. 1, issue 10, pp. 130–138 (in Russian).
9. Smirnov, B.M., Physics of Fractal Clusters,
10. Feder, J., Fractals,
11. Gusev, V., Living
Universe. Ch. IV-
12. Gusev, V.A. and Schulze-Makuch,
D., Genetic code: Lucky chance or fundamental law of nature? Physics
Life Rev., 2004, vol. 1, no. 3, pp. 202–229.
13. Gusev, V.A., Arithmetic
and algebra in the structure of genetic code, logic in the genome structure and
biochemical cycle of self-replication of living systems, Vestn.
VOGIS, 2005, vol. 9, no. 2,
pp. 153–161 (in Russian).
14. Carr, B. J. and Rees, M.Y. The anthropic principle and the structure of the physical
world, Nature, 1979, vol. 278, pp. 605–612.
15. Gorelik, G.E., Dimensionality
of Space,
16. Gorelik, G.E., Why Space
Is Three-Dimensional?
17. Ehrenfest, P. In what way does it become manifest in the fundamental laws of
physics that space has three dimensions? Proc. Amsterdam Acad.,
1917, vol. 20, pp. 200.
18. Dyatlov, V.L., Polarization
Model of Inhomogeneous Physical Vacuum,
19. Dmitriev, A.E., Dyatlov, V.L., and Gvozdarev, A.Yu., Unusual
Phenomena in Nature and Inhomogeneous Vacuum, Novosibirsk–Gorno-Altaisk–Biisk: Izd. BTPU im.
Shukshina,
2005 (in Russian).
20. Chernin, A.D., Dark energy and universal antigravitation, Usp. Fiz. Nauk, 2008, vol. 178, no. 3, pp. 267–300 (in Russian).
20a. Oparin, A.I., The
Origin of Life,
21. Strugatskie, A. and B., Billion
Years to the End of the Earth,
22. Astronomic Almanac (Constant Part), V.K. Abalkin, Ed.,
23. The System of Saturn, M.Ya.
Marov and V.M. Zharkov,
Eds.,
24. Gusev, V.A., Chemical
evolution in a thunderstorm cloud, Dokl.
Ross. Akad.
Nauk, 2002, vol. 385, no. 3, pp.
. 352–354 (in Russian).
25. Somsikov, V.M., To the origin of the physics of evolution, Zh. Problem Evol. Otkr. Sistem, 2006, vol. 2, issue 8, pp. 9–17 (in Russian).
2However, there
are some ideas how it is possible to conduct the experiments for in situ
real-time reproduction of the emergence of prebiotic structures
[24]. Also note here a pioneer work [25] that formalizes the description of nonequilibrium evolving systems.
3For convenience of scaling, the distances R
between satellites and planets are given in astronomic units
r = R/RE, where RE is the distance
from the Earth to Sun.